Размножение и развитие дождевых червей
Дождевые черви размножаются обычно в течение всего теплого времени года, в условиях умеренного климата — с весны до осени. Размножение прекращается только во время зимовки и засухи, когда черви перестают питаться и уходят в глубокие слои почвы.
Несмотря на наличие мужских и женских половых органов в одной особи, оплодотворение у червей обычно перекрестное. Имеются некоторые данные о возможности у них самооплодотворения (Evans a. Guild, 1947a). Однако скорее всего факты, относимые к самооплодотворению, следует толковать как случаи так называемого девственного размножения (см. стр. 59). Откладке яйцевых коконов обычно предшествует спаривание. Процесс спаривания прослежен у большого красного червя и навозного червя. Имеются некоторые наблюдения по спариванию длинного червя.
Рассмотрим процесс спаривания у навозного червя, где он хотя и протекает с некоторыми усложнениями вследствие расположения семеприемников на спинной стороне тела, но зато более полно известен и лучше изображен.
Спаривание может происходить на поверхности земли, но у навозного червя (а также у длинного червя) оно несомненно совершается и внутри грунта. Два червя сближаются ротовыми концами и, двигаясь навстречу друг другу, плотно соприкасаются брюшными сторонами в области передних 30—40 сегментов. Долгое время (до 1 часа) у обоих партнеров наблюдаются скольжение вперед в назад и обильное слизеотделение. Вокруг каждого червя образуется замкнутая трубка из слизи. Затем черви плотно слипаются брюшными сторонами таким образом, что сегменты 9-й—11-й, в которых расположены семеприемники одного партнера, располагаются против пояска другого партнера. Тогда железы, расположенные на брюшной стороне 9-го—11-го сегментов, обоих червей начинают усиленно выделять особое густое клейкое белое вещество, облекающее со всех сторон сегменты семеприемников одного партнера и сегменты пояска другого партнера (рис.
Передние концы двух дождевых червей, находящихся в копуляции
Железистые подушки 15-го сегмента, на котором находятся мужские половые отверстия, сильно набухают, наполняясь кровью, а на пространстве от 15-го сегмента до конца пояска появляются с каждой стороны тела глубокие складки — семенные бороздки. Образование семенных бороздок происходит действием особой мускулатуры, расположенной в стенке тела. В эти бороздки из мужских половых пор изливается семенная жидкость, которая движется назад по телу червя, к пояску. Достигнув уровня отверстий семеприемников другого партнера, семенная жидкость изливается туда.
Процесс спаривания может длиться несколько часов. Потревоженные черви (например, найденные в состоянии спаривания в грунте) расходятся не сразу, и за ними не трудно наблюдать, если предохранить их от сильного света и высыхания. После того как семеприемники обоих партнеров оказываются наполненными семенной жидкостью, черви расходятся и выползают из своих слизистых трубок. Такие копуляционные трубки бывают находимы в грунте. В них обычно остается избыточная часть семенной жидкости.
Некоторые виды дождевых червей лишены семеприемников. Как происходит спаривание у этих видов — пока не выяснено. Но известно, что отсутствие семеприемников заменяются у этих червей образованием сперматофоров. Так называются капли семенной жидкости, заключенные в засохшие слизистые выделения околощетинковых желез. Их находят прикрепленными в разных местах тела к кутикуле червей. По-видимому, это прикрепление совершается во время спаривания, причем сперматофоры, образованные каждым из червей, приклеиваются к кутикуле партнера при копуляции.
Яйца дождевых червей откладываются, как уже упомянуто, в яйцевых коконах. Раньше считалось, что коконы образуются во время спаривания. В недавнее время выяснено, что образование коконов — акт, не зависимый от спаривания.
Раньше считалось, что коконы образуются во время спаривания. В недавнее время выяснено, что образование коконов — акт, не зависимый от спаривания.
Процесс откладки кокона также начинается с образования слизистой трубки на участке тела примерно от 8-го до 35-го сегмента. Эта трубка захватывает все половые отверстия и поясок (если откладка кокона начинается сразу после спаривания, то, по-видимому, может использоваться копуляционная трубка). Железы пояска начинают усиленно функционировать, причем наибольшее значение имеет деятельность зернистых клеток. Крупнозернистые клетки выделяют вещество оболочки будущего кокона. Сначала оболочка кокона представляет собой не что иное, как совершенно прозрачную коллоидную пленку, расположенную под слизистой трубкой и охватывающую всю поверхность пояска. Под нею быстро накапливается густая опалесцирующая жидкость (выделение мелкозернистых клеток пояска) — содержимое кокона, представляющее собой питательный материал для яиц, откладываемых в кокон, так называемый белок яйцевых коконов. Процесс выделения жидкостей, образующих кокон, совершается поразительно быстро — в течение I —2 минут. Очевидно, это рефлекс, возможность которого обеспечивается наличием большого количества нервных окончаний в области пояска. Далее можно наблюдать, как по телу червя начинают пробегать характерные волны сокращения мускулатуры от пояска к ротовому концу, причем червь остается на месте. Образующийся кокон в виде белого толстого кольца, некоторое время находится у переднего края пояска, а затем начинает довольно быстро катиться к переднему концу тела. Мы употребляем слово «катиться» потому, что есть указания на «винтовой» характер этого движения образующегося кокона. При прохождении кокона через 12-й сегмент в его белок поступают из яйцеводов зрелые яйца, а при прохождении кокона через сегменты с порами семеприемников туда впрыскивается семенная жидкость, полученная при спаривании от другой особи. Разрезами через белок коконов установлено, что в момент образования около пояска белок не содержит сперматозоидов, последние появляются в нем только после прохождения формирующегося кокона через сегмент с семеприемниками (Grove, 1925; Grove a.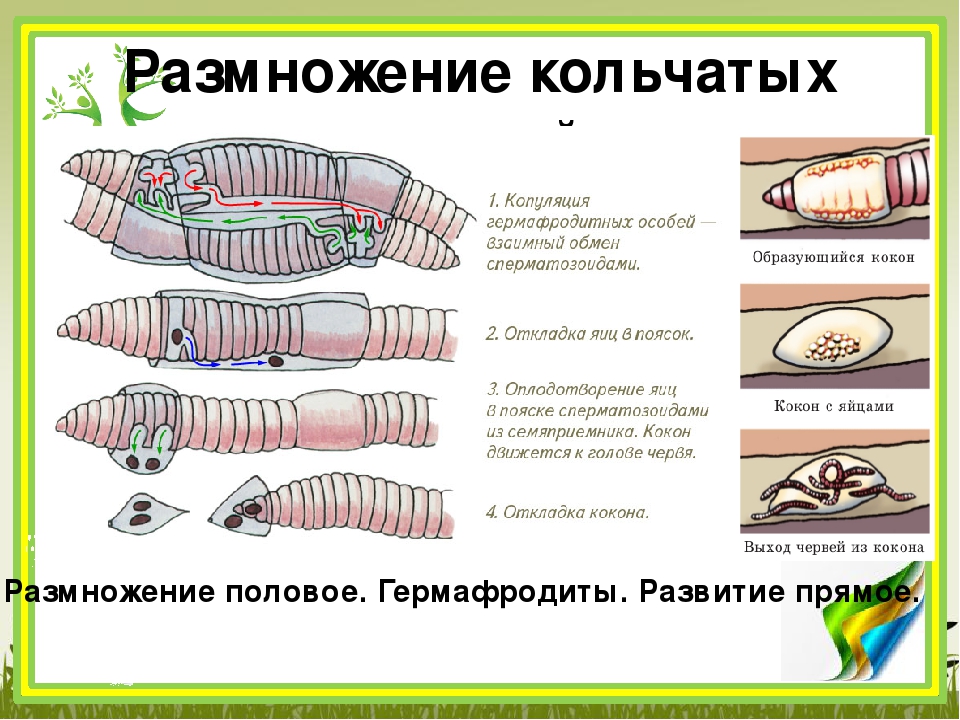
Только что отложенный кокон одет длинной слизистой трубкой, имеющей следы наружной сегментации червя (рис. 28). Оболочка кокона некоторое время остается бесцветной. При наблюдении за откладыванием кокона кажется, что с головы сбрасывается бесформенная масса мутной жидкости в слизистой трубке. На воздухе оболочка кокона быстро темнеет и твердеет; кокон приобретает вид овальной капсулы с двумя концами: передним, более широким и зубчатым, и задним, более тонким и обычно более длинным. Широкий конец трубки представляет собой канал, запечатанный, пробкой из ссохшегося белка. Слизистая трубка сохраняется на коконах очень недолго; ее наличие может служить указанием на то, что кокон имеет самое большее суточный возраст.
Только что отложенный яйцевой кокон навозного дождевого червя в слизистом чехле
Форма коконов у разных видов варьирует: она то более геометрически правильная, то угловатая; характер концевых трубочек также может быть различным. Окраска коконов бывает зелено-желтой, желтой, коричневой и бурой. Цвет характеризует не только вид, но и возраст кокона: с течением времени его поверхность несколько светлеет. Размеры коконов в длину колеблются от 2 до 6 мм.
Вопрос, как часто откладываются коконы одним червем, изучен недостаточно. Имеются сведения, что пашенный червь (Allolobophora caliginosa) откладывает коконы один раз в 7—10 дней. У навозного червя откладка коконов происходит несомненно чаще, но точных сведений об этом пока нет.
У большинства видов внутри кокона содержится не более 2—3 яиц, чаще всего — только одно. У навозного червя в коконе 6—20 яиц, но обычно развиваются далеко не все.
Оплодотворение яиц совершается в коконе уже после его откладки. Если вскрыть свежеотложенный кокон и выпустить его содержимое на предметное стекло, то под небольшим увеличением микроскопа в белке можно найти яйцо, окруженное сперматозоидами. Один из сперматозоидов проникает в яйцо. После этого у некоторых видов оболочка яйца (желточная оболочка) отстает от его поверхности. У других видов желточная оболочка бывает не видна.
Один из сперматозоидов проникает в яйцо. После этого у некоторых видов оболочка яйца (желточная оболочка) отстает от его поверхности. У других видов желточная оболочка бывает не видна.
Прежде чем говорить о развитии яиц, отметим, что, как установлено в последнее время, яйца ряда видов дождевых червей могут развиваться без оплодотворения (Muldal, 1950). Этот вариант полового размножения, называемый обычно девственным размножением, широко распространен среди беспозвоночных. Некоторые виды (например, большой красный червь — Lumbricus terrestris, длинный червь — Allolobophora longa) хотя и могут откладывать яйцевые коконы без предварительного опло-дотворенпя, но отложенные в них яйца не развиваются. У других (например, у пашенного червя — Allolobophora caliginosa) одни особи способны к девственному размножению, другие — неспособны. Однако, как правило, развитие без оплодотворения у этих видов в природе не происходит. Наконец, у водного дождевого червя (Eiseniella tetraedra), розового червя (Eisenia rosea) и у некоторых других видов, помимо особей, размножающихся с оплодотворением, есть особи, способные только к девственному размножению. Свойство яиц развиваться без оплодотворения облегчает червям заселение незанятых ими территорий, так как случайное попадание в новое место всего одного экземпляра может привести к возникновению впоследствии огромного числа особей.
При содержании в комнате развитие червей внутри коконов продолжается 2—3 недели. Но в естественных условиях оно обычно растягивается на значительно более длительные сроки, причем очень различные в зависимости от температурных и иных условий. Так, на Ротамстэдекой сельскохозяйственной станции в Англии развитие зародышей пашенного червя продолжалось (в зависимости от сезона) от 10 до 40 недель (Evans a. Guild, 1948).
Развитие начинается с того, что оплодотворенная яйцевая клетка многократно делится (рис. 29). В результате этого процесса дробления яйца возникает двуслойная пластинка, состоящая из клеток (рис. 30). Клетки будущего кишечника уже на этой стадии развития начинают поглощать окружающую их белковую массу, содержащуюся в коконе. Затем образуется своеобразная сферическая личинка, приспособленная для жизни только внутри яйцевого кокона. У нее имеется кишечник с ротовым и глоточным аппаратом, при помощи которого белковая жидкость накачивается в кишечник и, растягивая его, быстро увеличивает объем личинки во много раз (рис. 30). Нагнетание белковой жидкости в кишечник осуществляется действием ресничек, а обратный выход ее делается невозможным из-за особого клапана на границе между глоткой и кишечником. Заднепроходного отверстия у личинки нет, да оно и не нужно, так как белковая жидкость всасывается клетками кишечника без остатка.
30). Клетки будущего кишечника уже на этой стадии развития начинают поглощать окружающую их белковую массу, содержащуюся в коконе. Затем образуется своеобразная сферическая личинка, приспособленная для жизни только внутри яйцевого кокона. У нее имеется кишечник с ротовым и глоточным аппаратом, при помощи которого белковая жидкость накачивается в кишечник и, растягивая его, быстро увеличивает объем личинки во много раз (рис. 30). Нагнетание белковой жидкости в кишечник осуществляется действием ресничек, а обратный выход ее делается невозможным из-за особого клапана на границе между глоткой и кишечником. Заднепроходного отверстия у личинки нет, да оно и не нужно, так как белковая жидкость всасывается клетками кишечника без остатка.
Дробление яйца дождевого червя
Зародыш и личинка дождевого червя
Сферическая личинка вытягивается и мало-помалу приобретает червеобразную форму. На ней становятся видимыми границы между сегментами, размеры которых убывают спереди назад, так как образование новых сегментов происходит на заднем конце личинки. Спинная сторона формирующегося червя, развивается медленнее, чем брюшная. Поэтому вытянутые зародыши скоро приобретают характерную форму запятых, так как тело их сильно согнуто на спинную сторону, а толщина быстро убывает сзади наперед (рис. 31). По мере роста червячки постепенно приобретают цилиндрическую форму и у них формируются все органы, кроме полового аппарата, который появляется только у выросшего червя. Полное число сегментов, характерное для данного вида (от 80 до 250), образуется еще в коконе.
Формирующийся червь из яйцевого кокона
Издавна известно, что зародыши дождевых червей на стадиях дробления могут делиться пополам и давать начало однояйцевым близнецам. Имеются данные о том, что у Allolobophora caliginosa f. trapezoides в кокон откладывается всегда одно яйцо, но из него всегда выходят два червя, которые являются однояйцевыми близнецами (Omodeo, 1948; Omodeo a. Magalde, 1951). Таким образом, можно говорить о наличии у дождевых червей полиэмбрионии — особого способа размножения, свойственного некоторым насекомым (наездники) и некоторым низшим млекопитающим (армадилы).
Способность к активному движению черви получают очень рано. Сначала они двигаются внутри кокона при помощи ресничек. Затем возникают конвульсивные сокращения мускулатуры. Во вторую половину периода развития внутри кокона появляются типичные для них «червеобразные» перистальтические движения.
Белок яйцевого кокона по мере развития зародышей разжижается. В начале развития он представляет собой густую клейкую массу, а у некоторых видов имеет консистенцию густого желатина, так что его можно резать почти на куски. Еще на стадии сферической личинки можно заметить, что вокруг этой массы находится капля более жидкого белка. Это можно видеть в лупу через оболочку неповрежденного кокона. Количество жидкости в белке постепенно возрастает, и к концу развития червячки плавают в прозрачной, легко подвижной жидкости. Вряд ли можно сомневаться в том, что белок кокона постепенно переходит в раствор под действием ферментов, выделяемых телом зародыша. Таким образом, в кишечник попадает уже подготовленная к усвоению пища.
Оболочка коконов играет роль барьера, охраняющего содержимое от вторжения микробов и вредных веществ из почвы. Но она хорошо проницаема для воды и солей, что имеет большое значение для жизни зародышей (Светлов, 1928).
Вылупление червячков из кокона происходит при помощи их активных движений; они выползают передним концом вперед через более широкую трубочку кокона. Очень вероятно, что пробочка из засохшего белка, закупоривающая выход из этой трубочки, предварительно растворяется теми же ферментами, которые растворяют белок кокона во время развития. Выползание молодых червячков из кокона легко наблюдать, если положить в воду кокон, содержащий червячков, готовых к выходу. Такие коконы встречаются в земле нередко; их можно узнать по красному цвету крови в кровеносных сосудах молодых червячков, просвечивающей через прозрачную оболочку кокона. При погружении в воду червячки получают, очевидно, сильное раздражение, начинают беспокойно двигаться и вскоре один за другим выползают из кокона.
Вылупившиеся червячки имеют в длину 10—15 мм. Они обычно еще лишены пигмента. Почти сразу после выхода из кокона они начинают заглатывать землю и отыскивать пищу в виде разложившихся листочков, стебельков и пр.
О темпах роста червей после выхода их из яйцевого кокона наука пока располагает очень скудными сведениями. В естественных условиях большинство видов дождевых червей достигает половой зрелости примерно через год после выхода из кокона. У навозного червя этот срок, по-видимому, значительно меньше. В росте дождевых червей различают три периода: постнатальпый, период быстрого роста до образования пояска и, наконец, период медленного роста после образования пояска.
При старении вес дождевых червей падает (Michon, 1957). В общем, темпы развития очень сильно варьируют в зависимости от внешних условий. У трех видов рода Allolobophora и у розовой эйзении обнаружено, что в зимнее время происходит полная остановка развития молодых червей (Evans a. Guild, 1948). У пяти видов Allolobophora такого рода остановка в развитии, так называемая диапауза, может происходить и в теплое время года при понижении влажности почвы. Диапауза может длиться месяцами; при наступлении более благоприятных условий развитие продолжается (Michon, 1954, 1957).
Сведения о продолжительности жизни дождевых червей также довольно скудны. Известно, что в неволе навозные черви (Eisenia foetida) жили от 3/4 до 4% лет, большой красный червь (Lumbricus terrestris) — 5—6 лет, длинный червь (Allolobophora longa) — от 5 1/2 до 10% лет. Есть менее надежные сведения, что какой-то, ближе не определенный, дождевой червь прожил в неволе 15 лет. Эти факты говорят об относительно большом сроке их индивидуальной жизни, особенно если принять во внимание, что никто из содержавших червей длительное время в неволе не указывает на ухудшение их состояния или на какие-либо признаки старения. Таким образом, можно думать, что максимальная продолжительность жизни дождевых червей значительно превосходит приведенные выше цифры. Дождевым червям в довольно значительном масштабе свойственна способность к восстановлению утраченных частей тела, т. е. к регенерации. Головной конец восстанавливается при утрате спереди не более 15 сегментов, причем, сколько бы сегментов не было отрезано, восстанавливается только 4. Передняя отрезанная часть не восстанавливает задней и погибает; в редких случаях изолированные 5—6 головных сегментов восстанавливают на поверхности разреза вторую голову, т. е. получается уродливый нежизнеспособный организм, который также погибает. После отрезания заднего конца тела регенерация идет гораздо лучше. На задней раневой поверхности разрезанного червя всегда восстанавливается хвостовый конец, даже если оставлено только 30 передних сегментов (в редких случаях и при меньшем числе их). После заживления раны, недели через 1 1/2—2, появляется тоненький вырост (регенерат), размерами и цветом напоминающий червячка, только что вышедшего из яйцевого кокона. Он тем длиннее, чем большее число сегментов было отрезано. Регенерат растет в длину и в толщину, и месяца через 1 1/2 — 2 червь делается неотличимым от нормального.
Дождевым червям в довольно значительном масштабе свойственна способность к восстановлению утраченных частей тела, т. е. к регенерации. Головной конец восстанавливается при утрате спереди не более 15 сегментов, причем, сколько бы сегментов не было отрезано, восстанавливается только 4. Передняя отрезанная часть не восстанавливает задней и погибает; в редких случаях изолированные 5—6 головных сегментов восстанавливают на поверхности разреза вторую голову, т. е. получается уродливый нежизнеспособный организм, который также погибает. После отрезания заднего конца тела регенерация идет гораздо лучше. На задней раневой поверхности разрезанного червя всегда восстанавливается хвостовый конец, даже если оставлено только 30 передних сегментов (в редких случаях и при меньшем числе их). После заживления раны, недели через 1 1/2—2, появляется тоненький вырост (регенерат), размерами и цветом напоминающий червячка, только что вышедшего из яйцевого кокона. Он тем длиннее, чем большее число сегментов было отрезано. Регенерат растет в длину и в толщину, и месяца через 1 1/2 — 2 червь делается неотличимым от нормального.
При разрезании червя пополам передняя половина восстанавливает хвостовой конец; головной конец на задней половине не восстанавливается. Однако и здесь возможна патологическая регенерация: па переднем конце задней половины в некоторых случаях, после длительной паузы, появляется регенерат, но оказывается, что это не регенерат головы, а второй хвост. Такие уродливые формы гибнут (рис. 32).
Регенерация у дождевого червя
Отметим, что хотя способность к регенерации у дождевых червей гораздо лучше выражена, чем у позвоночных, так как они могут восстанавливать довольно крупные отделы тела, но ее не следует переоценивать. По сравнению с низшими формами малощетинковых червей люмбрициды восстанавливают утраченные части в значительно меньших масштабах и гораздо медленнее.
Культивирование (размножение) дождевых червей на садовом участке
В саду, огороде, где-нибудь в тени деревьев или под навесом, в сарае, подвале и т. д., в ящике, в старой ванне или прямо на земле необходимо положить слой компоста толщиной 40-50 см в виде насыпной грядки. Разровняйте и хорошо увлажните. Влажность достаточная, если из комка компоста, зажатого в кулак, появятся 1-2 капли влаги. Размер червятника первоначально не должен быть большим (достаточно 2м2). Хорошо увлажнённый субстрат в культиваторе закройте старой мешковиной (или чёрной перфорированной плёнкой, или соломой). Влажный субстрат должен выстояться 5-7 суток. За это время его надо периодически увлажнять. Это необходимо для удаления остатков аммиака и растворения кристалликов солей удобрений, которые могут причинить червям некоторый вред.
д., в ящике, в старой ванне или прямо на земле необходимо положить слой компоста толщиной 40-50 см в виде насыпной грядки. Разровняйте и хорошо увлажните. Влажность достаточная, если из комка компоста, зажатого в кулак, появятся 1-2 капли влаги. Размер червятника первоначально не должен быть большим (достаточно 2м2). Хорошо увлажнённый субстрат в культиваторе закройте старой мешковиной (или чёрной перфорированной плёнкой, или соломой). Влажный субстрат должен выстояться 5-7 суток. За это время его надо периодически увлажнять. Это необходимо для удаления остатков аммиака и растворения кристалликов солей удобрений, которые могут причинить червям некоторый вред.Через 5-7 дней в центре каждого квадратного метра культиватора сделайте лопатой ямку, как для посадки картофеля, и в неё опрокиньте ведро с подготовленными червями. Выровняйте поверхность и закройте воздухопроницаемым материалом (мешковиной, соломой и т. п.). Через сутки увлажните субстрат культиватора. В сухую и жаркую погоду необходимо поливать его водой не чаще, чем огурцы.
Такой способ заселения культиватора червями связан с тем, что червям новый субстрат может показаться «невкусным», и тогда они будут некоторое время отсиживаться в своём родном субстрате. Но голод заставит их снова и снова пробовать новый для них субстрат. Так постепенно они привыкнут к новому корму и новому дому.
Через неделю после заселения посмотрите, переходят ли черви в новый субстрат, этого срока достаточно для его освоения. Если поверхность червей чистая, а сами они подвижны — это свидетельство их благополучия. Если они вялые, не активные, не пытаются прятаться от света — это признаки их тяжёлого поражения различными пестицидами из нового для них корма. В этом случае нужно найти новую популяцию (партию) червей из другой навозной кучи и посадить их в культиватор. Возможно придётся создавать новый компост из другого источника органики. Но такая необходимость возникает крайне редко.
Если черви чувствуют себя хорошо в новом доме, то на 3-4 недели их нужно оставить в покое. Единственное, в чем они нуждаются в течение этого времени, так это во влаге. Периодически поливайте грядку-культиватор водой, температура которой должна быть равной температуре окружающей среды. Слишком холодная или слишком тёплая вода вызывает у червей так называемую стрессовую реакцию (испуг, шок), и они перестают хорошо питаться и размножаться. Вода для полива культиватора с червями должна храниться в какой-либо ёмкости и выстаиваться не менее суток. За это время она прогреется до нужной температуры, и из неё выветрится хлор (если вода из городского водопровода).
Единственное, в чем они нуждаются в течение этого времени, так это во влаге. Периодически поливайте грядку-культиватор водой, температура которой должна быть равной температуре окружающей среды. Слишком холодная или слишком тёплая вода вызывает у червей так называемую стрессовую реакцию (испуг, шок), и они перестают хорошо питаться и размножаться. Вода для полива культиватора с червями должна храниться в какой-либо ёмкости и выстаиваться не менее суток. За это время она прогреется до нужной температуры, и из неё выветрится хлор (если вода из городского водопровода).
После адаптации (приспособления) к новым условиям вся деятельность червей будет направлена на откладку коконов (капсул, величиной в половину рисового зерна лимоноподобной формы, жёлтых, с мягкой, но прочной оболочкой). В каждом коконе от 3 до 21 зародыша. Каждый червь откладывает по одному кокону за 5-7 дней в течение 12-18 недель. Из отложенных коконов через 15.. .20 суток (в зависимости от температуры субстрата) появляются маленькие новорождённые червята, тонкие, как нитки, длиной около 4.. .6 мм, с красным хорошо видным спинальным кровеносным сосудом. Это их отличительный признак от нематоды — малых белых червей, которые не имеют красного кровеносного сосуда. Червята подрастают быстро и за 10-12 недель увеличивают свою массу с 1 до 250-500 мг. Последние коконы (в районе Подмосковья) откладываются червями в конце июля, а последние червята появляются на свет до 20 августа.
Червята подрастают быстро и в подавляющем большинстве становятся вполне взрослыми к октябрю. В течение летнего сезона количество червей и их общая живая масса в культиваторе увеличивается в 20-50 раз.
Для их размножения и роста требуется много пищи. Поэтому в червятник (грядку-культиватор) необходимо периодически добавлять корм в виде компоста, наслаивая его по 15-20 см через каждые 2-3 недели, начиная с первых чисел июня. Последнее кормление червей необходимо провести в конце октября или даже в начале ноября до наступления морозов. При понижении температуры черви снижают свою активность: движения их замедляются, при температуре 6°С черви перестают питаться, а при 4° С освобождают свой пищеварительный тракт от остатков пищи и начинают впадать в состояние анабиоза (зимней «спячки»). С наступлением морозов они замерзают. Но это для них не опасно. Они пережили все ледниковые периоды. С наступлением весны они оживают и начинают новый активный период своей жизни.
При понижении температуры черви снижают свою активность: движения их замедляются, при температуре 6°С черви перестают питаться, а при 4° С освобождают свой пищеварительный тракт от остатков пищи и начинают впадать в состояние анабиоза (зимней «спячки»). С наступлением морозов они замерзают. Но это для них не опасно. Они пережили все ледниковые периоды. С наступлением весны они оживают и начинают новый активный период своей жизни.
За время летнего культивирования приходится делать 7-8 наслоений компостов. По мере их поедания червями они уплотняются, но всё же грядка-культиватор становится всё выше и выше. Высота её осенью может достигать 0,6 м. Она легко продувается ветром, в ней труднее поддерживать необходимую влажность. Исходя из этого, боковые поверхности её рекомендуется заделать досками (в виде ящика).
В ходе культивирования за летний период в червятнике будет переработано с помощью червей более 1 т компоста (50% влажности) на каждом квадратном метре его площади.
Черви к осени располагаются в культиваторе в основном в верхнем (пищевом) слое, тогда как нижний слой заселён червями слабо — он используется ими как «санитарный блок» для испражнений. Толщина верхнего слоя около 20 см. Нижний слой состоит в основном из копролитов и представляет собой гумусное органическое удобрение (биогумус, червекомпост), сырое и нуждающееся в подработке. Это тот продукт, ради которого и осуществляется культивирование червей.
Дождевые черви бывают гигантскими и способны спариваться часами
- Мэтт Уокер
- BBC Earth
Автор фото, ARCO/naturepl.com
Обозреватель BBC Earth рассказывает об удивительных существах, живущих группами и совершающих инженерные подвиги.
Дождевой червь по имени Дэйв взбудоражил британскую научную общественность. А все потому, что длина Дэйва 40 см, что делает его и ее (мы объясним это позже) самым длинным дождевым червем, найденным на территории Великобритании.
Но — хоть и это может прозвучать неожиданно — о дождевых червях можно рассказать еще много чего интересного.
Вот пять удивительных фактов о них.
1. Дождевые черви достигают невероятных размеров
Автор фото, blickwinkel / Alamy Stock Photo
Подпись к фото,Гигантский Megascolides australis
Допустим, дождевой червь Дэйв действительно удивил граждан Великобритании, включая исследователей из лондонского Музея естественной истории, измерявших его.
Однако — по сравнению со своими сородичами из других уголков мира — с длиной в 40 сантиметров он просто кроха, если учесть, что некоторые виды достигают 1 м.
Измерить длину дождевого червя довольно непросто, так как его мягкое тело естественным образом вытягивается и сокращается, и особь можно случайно растянуть.
Тем не менее самым крупным дождевым червем в мире может быть Megascolides australis, обитающий в Австралии. Это редкий вид, встречающийся всего в пяти регионах этой страны.
Согласно Энциклопедии находящихся под угрозой видов, представители Megascolides australis достигают 2 м в длину при диаметре 4 см.
Другой вид, африканский гигантский дождевой червь (Microchaetus rappi), тоже, как говорят, может похвастаться длиной в 2 м и более.
2. Гигантские дождевые черви создают загадочные холмы
Автор фото, Deilphine Renard
Подпись к фото,Снимок «суралес» с беспилотника
Известно, что черви способны перепахивать землю, но в Южной Америке один из видов дождевых червей создает необычные, но очень красивые холмики.
«Суралес» — это ландшафт, состоящий из зеленых бугорков и глубоких ям, образующих замысловатые, но в то же время правильные узоры.
Они покрывают множество квадратных километров, однако эти удивительные холмы были созданы не кем иным, как обычными дождевыми червями длиной до 1 м.
Способ, при помощи которого дождевые черви строят свои бугорки, несколько странный.
За многие десятилетия черви вида Andiorrhinus приобрели строгие привычки в питании. Их трапеза проходит в затопленных ямах, и свои экскременты они оставляют там же.
Со временем их экскременты накапливаются и образуют огромные бугры до 5 м в ширину и 2 м в высоту. Об этом открытии ученые сообщили совсем недавно — в мае 2016 года.
3. Дождевые черви способны спариваться часами
Автор фото, Robin Chittenden/naturepl.com
Подпись к фото,Обыкновенные земляные черви (Lumbricus terrestris) в процессе спаривания
Что касается сексуальной выносливости, то дождевые черви могут легко превзойти другие виды, в том числе человека.
Спаривание у обыкновенных дождевых червей (Lumbricus terrestris) может длиться более трех часов подряд.
Исследование 1997 года показало, что во время ухаживания дождевые черви залезают в норы друг к другу, иногда до 17 раз подряд.
Если этого вам недостаточно для подтверждения их стойкости и запасов энергии, вот еще один факт: каждый половой акт длится от 69 до 200 минут. Ученым это известно, так как они сняли сам процесс на видео.
4. У них проблемы с отцовством (и материнством)
Автор фото, Kim Taylor/naturepl.com
Подпись к фото,И папа, и мама… Отсюда все проблемы
Дождевые черви — гермафродиты, то есть у каждой особи имеются и мужские, и женские половые органы.
Во время спаривания они борются за то, чтобы передать друг другу семенную жидкость и оплодотворить яйцеклетки друг друга.
Это означает, что Дэйв мог быть и отцом, и матерью многочисленных отпрысков, передав каждому из них половину своих генов.
Но на этом трудности не заканчиваются.
В 2013 году ученые впервые в истории провели тесты на отцовство в отношении дождевых червей. Они обнаружили, что у них зачастую бывает несколько партнеров, и шансы получить потомство выше всего у первого и третьего червя, участвовавших в спаривании с конкретным партнером.
Они обнаружили, что у них зачастую бывает несколько партнеров, и шансы получить потомство выше всего у первого и третьего червя, участвовавших в спаривании с конкретным партнером.
Вероятно, второй червь терпит неудачу потому, что его семенная жидкость не достигает семяприемника, уже заполненного первым партнером, и затем смывается семенной жидкостью третьего.
Более того, в то время как некоторые дождевые черви спариваются очень долго, другие виды вообще отказались от спаривания.
У некоторых тайваньских горных дождевых червей (Amynthas catenus) в процессе эволюции исчезли гениталии, и потомство они производят при помощи девственного размножения, или партеногенеза.
Это означает, что они передают своим отпрыскам 100 % своих генов, создавая собственных клонов, что делает их и матерью, и отцом каждому маленькому червю.
5. Они образуют стада, подобно коровам
Автор фото, ARCO/naturepl.com
Подпись к фото,Стадо дождевых червей
А еще они принимают групповые решения. По данным исследования, опубликованного в 2010 году, дождевые черви способны общаться между собой и влиять на поведение друг друга при помощи прикосновений.
Это дает им возможность коллективно решать, в каком направлении будет двигаться все стадо.
Это удивительное поведение червей вида Eisenia fetida впервые позволило доказать, что некоторые кольчатые черви действительно способны образовывать активные группы.
Из этого открытия следует, что дождевые черви — социальные животные, и их групповое поведение схоже с тем, что демонстрируют другие виды, ведущие стадный образ жизни.
Так что в следующий раз, когда вы возьмете в руки червя и бросите его в компостную кучу, вспомните об этом. Возможно, вы разлучаете его с друзьями.
Размножение и развитие дождевых червей
Дождевые черви размножаются обычно в течение всего теплого времени года, в условиях умеренного климата — с весны до осени. Размножение прекращается только во время зимовки и засухи, когда черви перестают питаться и уходят в глубокие слои почвы.
Размножение прекращается только во время зимовки и засухи, когда черви перестают питаться и уходят в глубокие слои почвы.
Несмотря на наличие мужских и женских половых органов в одной особи, оплодотворение у червей обычно перекрестное. Имеются некоторые данные о возможности у них самооплодотворения (Evans a. Guild, 1947а). Однако скорее всего факты, относимые к самооплодотворению, следует толковать как случаи так называемого девственного размножения. Откладке яйцевых коконов обычно предшествует спаривание. Процесс спаривания прослежен у большого красного червя и навозного червя. Имеются некоторые наблюдения по спариванию длинного червя.
Рассмотрим процесс спаривания у навозного червя, где он хотя и протекает с некоторыми усложнениями вследствие расположения семеприемников на спинной стороне тела, но зато более полно известен и лучше изображен.
Спаривание может происходить на поверхности земли, но у навозного червя (а также у длинного червя) оно, несомненно совершается и внутри грунта. Два червя сближаются ротовыми концами и, двигаясь навстречу друг другу, плотно соприкасаются брюшными сторонами в области передних 30 — 40 сегментов. Долгое время (до 1 часа) у обоих партнеров наблюдаются скольжение вперед в назад и обильное слизеотделение. Вокруг каждого червя образуется замкнутая трубка из слизи. Затем черви плотно слипаются брюшными сторонами таким образом, что сегменты 9-й — 11-й, в которых расположены семяприёмники одного партнера, располагаются против пояска другого партнера. Тогда железы, расположенные на брюшной стороне 9-го — 11-го сегментов, обоих червей начинают усиленно выделять особое густое клейкое белое вещество, облекающее со всех сторон сегменты семяприёмников одного партнера и сегменты пояска другого партнера (рисунок 27). Сегменты пояска плотно присасываются к телу партнера валиками зрелости, и между последними образуется желобок, в который погружается область тела 9-го — 11-го сегментов другого червя. В этих местах тело обоих червей перехвачено как бы двумя повязками.
Железистые подушки 15-го сегмента, на котором находятся мужские половые отверстия, сильно набухают, наполняясь кровью, а на пространстве от 15-го сегмента до конца пояска появляются с каждой стороны тела глубокие складки — семенные бороздки. Образование семенных бороздок происходит действием особой мускулатуры, расположенной в стенке тела. В эти бороздки из мужских половых пор изливается семенная жидкость, которая движется назад по телу червя, к пояску. Достигнув уровня отверстий семяприёмников другого партнера, семенная жидкость изливается туда.
Рисунок. 27. Передние концы двух дождевых червей, находящихся в копуляции. (По Каули).
Путь от мужских отверстий (1) до семеприемников (2) обозначен стрелками.
Процесс спаривания может длиться несколько часов. Потревоженные черви (например, найденные в состоянии спаривания в грунте) расходятся не сразу, и за ними нетрудно наблюдать, если предохранить их от сильного света и высыхания. После того как семяприёмники обоих партнеров оказываются наполненными семенной жидкостью, черви расходятся и выползают из .своих слизистых трубок. Такие копуляционные трубки бывают находимы в грунте. В них обычно остается избыточная часть семенной жидкости.
Некоторые виды дождевых червей лишены семяприёмников. Как происходит спаривание у этих видов — пока не выяснено. Но известно, что отсутствие семяприёмников заменяются у этих червей образованием сперматофоров. Так называются капли семенной жидкости, заключенные в засохшие слизистые выделения околощетпнковых желез. Их находят прикрепленными в разных местах тела к кутикуле червей. По-видимому, это прикрепление совершается во время спаривания, причем сперматофоры, образованные каждым из червей, приклеиваются к кутикуле партнера при копуляции.
Яйца дождевых червей откладываются, как уже упомянуто, в яйцевых коконах. Раньше считалось, что коконы образуются во время спаривания. В недавнее время выяснено, что образование коконов — акт, не зависимый от спаривания.
Процесс откладки кокона также начинается с образования слизистой трубки на участке тела примерно от 8-го до 35-го сегмента.
Эта трубка захватывает все половые отверстия и поясок (если откладка кокона начинается сразу после спаривания, то, по-видимому, может использоваться копуляционная трубка). Железы пояска начинают усиленно функционировать, причем наибольшее значение имеет деятельность зернистых клеток. Крупнозернистые клетки выделяют вещество оболочки будущего кокона. Сначала оболочка кокона представляет собой не что иное, как совершенно прозрачную коллоидную пленку, расположенную под слизистой трубкой и охватывающую всю поверхность пояска. Под нею быстро накапливается густая опалесцирующая жидкость (выделение мелкозернистых клеток пояска) — содержимое кокона, представляющее собой питательный материал для яиц, откладываемых в кокон, так называемый белок яйцевых коконов.
Процесс выделения жидкостей, образующих кокон, совершается поразительно быстро — в течение I — 2 минут.
Очевидно, это рефлекс, возможность которого обеспечивается наличием большого количества нервных окончаний в области пояска. Далее можно наблюдать, как по телу червя начинают пробегать характерные волны сокращения мускулатуры от пояска к ротовому концу, причем червь остается на месте. Образующийся кокон в виде белого толстого кольца, некоторое время находится у переднего края пояска, а затем начинает довольно быстро катиться к переднему концу тела. Мы употребляем слово «катиться» потому, что есть указания на «винтовой» характер этого движения образующегося кокона. При прохождении кокона через 12-й сегмент в его белок поступают из яйцеводов зрелые яйца, а при прохождении кокона через сегменты с порами семяприёмников туда впрыскивается семенная жидкость, полученная при спаривании от другой особи. Разрезами через белок коконов установлено, что в момент образования около пояска белок не содержит сперматозоидов, последние появляются в нем только после прохождения формирую Рисунок. 28. Только щегося кокона через сегмент с семепри что отложен-емниками. Затем кокон сбрасывается кокон навозного дождевого через голову. Весь процесс образования червя в слизи-кокона занимает 3 — 4 минуты.
28. Только щегося кокона через сегмент с семепри что отложен-емниками. Затем кокон сбрасывается кокон навозного дождевого через голову. Весь процесс образования червя в слизи-кокона занимает 3 — 4 минуты.
(Из Фюллера). длинной слизистой трубкой, имеющей следы наружной сегментации червя (рисунок 28).
Оболочка кокона некоторое время остается бесцветной. При наблюдении за откладыванием кокона кажется, что с головы сбрасывается бесформенная масса мутной жидкости в слизистой трубке. На воздухе оболочка кокоца быстро темнеет и твердеет; кокон приобретает вид овальной капсулы с двумя концами: передним, более широким и зубчатым, и задним, более тонким и обычно более длинным. Широкий конец трубки представляет собой канал, запечатанный, пробкой из ссохшегося белка. Слизистая трубка сохраняется на коконах очень недолго; ее наличие может служить указанием на то, что кокон имеет самое большее суточный возраст.
Форма коконов у разных видов варьирует: она то более геометрически правильная, то угловатая; характер концевых трубочек также может быть различным.
Окраска коконов бывает зелено-желтой, желтой, коричневой и бурой. Цвет характеризует не только вид, но и возраст кокона: с течением времени его поверхность несколько светлеет. Размеры коконов в длину колеблются от 2 до 6 мм.
Вопрос, как часто откладываются коконы одним червем, изучен недостаточно. Имеются сведения, что пашенный червь (Allolobophora caliginosa) откладывает коконы один раз в 7 — 10 дней. У навозного червя откладка коконов происходит несомненно чаще, но точных сведений об этом пока нет.
У большинства видов внутри кокона содержится не более 2 — 3 яиц, чаще всего — только одно. У навозного червя в коконе 6 — 20 яиц, но обычно развиваются далеко не все.
Оплодотворение яиц совершается в коконе уже после его откладкии. Если вскрыть свежеуложенный кокон и выпустить его содержимое на предметное стекло, то под небольшим увеличением микроскопа в белке можно найти яйцо, окруженное сперматозоидами. Один из сперматозоидов проникает в яйцо. После этого у некоторых видов оболочка яйца (желточная оболочка) отстает от его поверхности. У других видов желточная оболочка бывает не видна.
Один из сперматозоидов проникает в яйцо. После этого у некоторых видов оболочка яйца (желточная оболочка) отстает от его поверхности. У других видов желточная оболочка бывает не видна.
Прежде чем говорить о развитии яиц, отметим, что, как установлено в последнее время, яйца ряда видов дождевых червей могут развиваться без оплодотворения (Muldal, 1950). Этот вариант полового размножения, называемый обычно девственным размножением, широко распространен среди беспозвоночных. Некоторые виды (например, большой красный червь — Lumbricus terrestris, длинный червь — Allolobophora longa) хотя и могут откладывать яйцевые коконы без предварительного оплодотворения, но отложенные в них яйца не развиваются. У других (например, у пашенного червя — Allolobophora caliginosa) одни особи способны к девственному размножению, другие — неспособны. Однако, как правило, развитие без оплодотворения у этих видов в природе не происходит. Наконец, у водного дождевого червя (Eiseniella tetraedra), розового червя (Eisenia rosea) и у некоторых других видов, помимо особей, размножающихся с оплодотворением, есть особи, способные только к девственному размножению. Свойство яиц развиваться без оплодотворения облегчает червям заселение незанятых ими территорий, так как случайное попадание в новое место всего одного экземпляра может привести к возникновению впоследствии огромного числа особей.
При содержании в комнате развитие червей внутри коконов продолжается 2 — 3 недели. Но в естественных условиях оно обычно растягивается на значительно более длительные сроки, причем очень различные в зависимости от температурных и иных условий. Так, на Ротамстэдской сельскохозяйственной станции в Англии развитие зародышей пашенного червя продолжалось (в зависимости от сезона) от 10 до 40 недель (Evans a. Guild, 1948).
Рисунок. 29. Дробление яйца дождевого червя. (По П. Г. Светлову).
А — яйцо до дробления; Б — стадия 2 клеток; В — стадия 4 клеток; Г — стадия 12 клеток; Л — более поздняя стадия дробления яйца.
Пунктированы клетки, образующие стенку кишечника.
Развитие начинается с того, что оплодотворенная яйцевая клетка многократно делится.
В результате этого процесса дробления яйца возникает двуслойная пластинка, состоящая из клеток (рисунок 30). Клетки будущего кишечника уже на этой стадии развития начинают поглощать окружающую их белковую массу, содержащуюся в коконе. Затем образуется своеобразная сферическая личинка, приспособленная для жизни только внутри яйцевого кокона. У нее имеется кишечник с ротовым и глоточным аппаратом, при помощи которого белковая жидкость накачивается в кишечник и, растягивая его, быстро увеличивает объем личинки во много раз (рисунок 30). Нагнетание белковой жидкости в кишечник осуществляется действием ресничек, а обратный выход ее делается невозможным из-за особого клапана на границе между глоткой и кишечником. Заднепроходного отверстия у личинки нет, да оно и не нужно, так как белковая жидкость всасывается клетками кишечника без остатка.
Рисунок. 30. Зародыш и личинка дождевого червя. (По П. Г. Светлову).
А — плоский зародыш; нижний слой пластинки» состоящий ив крупных клеток, образует впоследствии стенку кишечника; Б — личинка, питающаяся белком кокона: слева — ротовое отверстие, внутри личинки — полость кишечника.
Сферическая личинка вытягивается и мало-помалу приобретает червеобразную форму. На ней становятся видимыми границы между сегментами, размеры которых убывают спереди назад, так как образование новых сегментов происходит на заднем конце личинки. Спинная сторона формирующегося червя, развивается медленнее, чем брюшная. Поэтому вытянутые зародыши скоро приобретают характерную форму запятых, так как тело их сильно согнуто на спинную сторону, а толщина быстро убывает сзади наперед (рисунок 31). По мере роста червячки постепенно приобретают цилиндрическую форму и у них формируются все органы, кроме полового аппарата, который появляется только у выросшего червя. Полное число сегментов, характерное для данного вида (от 80 до 250), образуется еще в коконе.
Полное число сегментов, характерное для данного вида (от 80 до 250), образуется еще в коконе.
Издавна известно, что зародыши дождевых червей на стадиях дробления могут делиться пополам и давать начало однояйцевым близнецам. Имеются данные о том, что у Allolobophora caliginosa f. trapezoides u кокон откладывается всегда одно яйцо, но из него всегда выходят два червя, которые являются однояйцевыми близнецами (Omodeo, 1948; Omodeo a. Magalde, 1951). Таким образом, можно говорить о наличии у дождевых червей полиэмбрионии — особого способа размножения, свойственного некоторым насекомым (наездники) и некоторым низшим млекопитающим (армадилы).
Способность к активному движению черви получают очень рано. Сначала они двигаются внутри кокона при помощи ресничек. Затем возникают конвульсивные сокращения мускулатуры. Во вторую половину периода развития внутри кокона появляются типичные для них «червеобразные» перистальтические движения.
Рисунок. 31. Формирующийся чернь из яйцевого кокона. (По Фюллеру).
Слева — передний конец тела с ротовым отверстием и головной лопастью; справа — задний конец тела с формирующимися новыми сегментами.
Белок яйцевого кокона по мере развития зародышей разжижается.
В начале развития он представляет собой густую клейкую массу, а у некоторых видов имеет консистенцию густого желатина, так что его можно резать почти на куски. Еще на стадии сферической личинки можно заметить, что вокруг этой массы находится капля более жидкого белка. Это можно видеть в лупу через оболочку неповрежденного кокона. Количество жидкости в белке постепенно возрастает, и к концу развития червячки плавают в прозрачной, легко подвижной жидкости. Вряд ли можно сомневаться в том, что белок кокона постепенно переходит в раствор под действием ферментов, выделяемых телом зародыша. Таким образом, в кишечник попадает уже подготовленная к усвоению пища.
Оболочка коконов играет роль барьера, охраняющего содержимое от вторжения микробов и вредных веществ из почвы. Но она хорошо проницаема для воды и солей, что имеет большое значение для жизни зародышей (Светлов, 1928).
Но она хорошо проницаема для воды и солей, что имеет большое значение для жизни зародышей (Светлов, 1928).
Вылупление червячков из кокона происходит при помощи их активных движений; они выползают передним концом вперед через более широкую трубочку кокона. Очень вероятно, что пробочка из засохшего белка, закупоривающая выход из этой трубочки, предварительно растворяется теми же ферментами, которые растворяют белок кокона во время развития. Выползание молодых червячков из кокона легко наблюдать, если положить в воду кокон, содержащий червячков, готовых к выходу. Такие коконы встречаются в земле нередко; их можно узнать по красному цвету крови в кровеносных сосудах молодых червячков, просвечивающей через прозрачную оболочку кокона. При погружении в воду червячки получают, очевидно, сильное раздражение, начинают беспокойно двигаться и вскоре один за другим выползают из кокона.
Вылупившиеся червячки имеют в длину 10 — 15 мм. Они обычно еще лишены пигмента. Почти сразу после выхода из кокона они начинают заглатывать землю и отыскивать пищу в виде разложившихся листочков, стебельков и пр.
О темпах роста червей после выхода их из яйцевого кокона наука пока располагает очень скудными сведениями.
В естественных условиях большинство видов дождевых червей достигает половой зрелости примерно через год после выхода из кокона. У навозного червя этот срок, по-видимому, значительно меньше. В росте дождевых червей различают три периода: постнатальный, период быстрого роста до образования пояска и, наконец, период медленного роста после образования пояска.
При старении вес дождевых червей падает (Michon, 1957). В общем, темпы развития очень сильно варьируют в зависимости от внешних условий. У трех видов рода Allolobophora и у розовой эйзении обнаружено, что в зимнее время происходит полная остановка развития молодых червей (Evans a. Guild, 1948). У пяти видов Allolobophora такого рода остановка в развитии, так называемая диа-пауза, может происходить и в теплое время года при понижении влажности почвы. Диапауза может длиться месяцами; при наступлении более благоприятных условий развитие продолжается (Michon, 1954, 1957).
Диапауза может длиться месяцами; при наступлении более благоприятных условий развитие продолжается (Michon, 1954, 1957).
Сведения о продолжительности жизни дождевых червей также довольно скудны. Известно, что в неволе навозные черви (Eisenia foetida) жили от 3/4 до 4% лет, большой красный червь (Lumbricus ter- restris) — 5 — 6 лет, длинный червь (Allolobophora longa) — от 5% до 10% лет. Есть менее надежные сведения, что какой-то, ближе не определенный, дождевой червь прожил в неволе 15 лет. Эти факты говорят об относительно большом сроке их индивидуальной жизни, особенно если принять во внимание, что никто из содержавших червей длительное время в неволе не указывает на ухудшение их состояния или на какие-либо признаки старения. Таким образом, можно думать, что максимальная продолжительность жизни дождевых червей значительно превосходит приведенные выше цифры.
Рисунок. 32. Регенерация у дождевого червя. Из середины тела было вырезано 13 сегментов. Сзади восстановилась хвостовая часть. Спереди образовались два хвостовых конца..
Дождевым червям в довольно значительном масштабе свойственна способность к восстановлению утраченных частей тела, т. е. к регенерации. Головной конец восстанавливается при утрате спереди не более 15 сегментов, причем, сколько бы сегментог не было отрезано, восстанавливается только 4. Передняя отрезанная часть не восстанавливает задней и погибает; в редких случаях изолированные 5 — б головных сегментов восстанавливают на поверхности разреза вторую голову, т. е. получается уродливый нежизнеспособный организм, который также погибает. После отрезания заднего конца тела регенерация идет гораздо лучше. На задней раневой поверхности разрезанного червя всегда восстанавливается хвостовой конец, даже если оставлено только 30 передних сегментов (в редких случаях и при меньшем числе их). После заживления раны, недели через 1/4 — 2, появляется тоненький вырост (регенерат), размерами и цветом напоминающий червячка, только что вышедшего из яйцевого кокона. Он тем длиннее, чем большее число сегментов было отрезано. Регенерат растет в длину и в толщину, и месяца через 2 червь делается неотличимым от нормального.
Он тем длиннее, чем большее число сегментов было отрезано. Регенерат растет в длину и в толщину, и месяца через 2 червь делается неотличимым от нормального.
При разрезании червя пополам передняя половина восстанавливает хвостовой конец; головной конец на задней половине не восстанавливается. Однако и здесь возможна патологическая регенерация: па переднем конце задней половины в некоторых случаях, после длительной паузы, появляется регенерат, но оказывается, что это не регенерат головы, а второй хвост. Такие уродливые формы гибнут (рисунок 32).
Отметим, что хотя способность к регенерации у дождевых червей гораздо лучше выражена, чем у позвоночных, так как они могут восстанавливать довольно крупные отделы тела, но ее не следует переоценивать. По сравнению с низшими формами малощетинковых червей люмбрициды восстанавливают утраченные части в значительно меньших масштабах и гораздо медленнее.
Поделиться:
Скопировать ссылку
Также рекомендуем прочитать:
Способы размножения дождевых червей | Ворми Ферма
На чтение 5 мин. Просмотров 3.1k. Опубликовано Обновлено
Земляные черви – уязвимые животные, но, в то же время, они обладают исключительными механизмами выживания, благодаря чему так широко распространены. Один из механизмов – размножение дождевого червя. Они обеспечивают высокую сохранность будущему потомству, появляются на свет в виде полноценных червей, минуя стадию личинки, и с легкостью размножаются в неволе
Как рождаются земляные червиПоявляются черви из яиц, которые заключены в коконы с прочной оболочкой. На вид это овальная капсула диаметром 2-4 мм, содержащая от 1 до 20 яиц (зависит от вида беспозвоночных).
Естественно
оплодотворяются и развиваются не все из яиц
Способы размножение червей:
Бесполое размножение дождевого червяТак как органы и системы у червей распределены равномерно по всем сегментам туловища, то при делении обе половины сохранят жизненно-важные функции. Но при условии, что с каждой стороны осталось достаточное число сегментов. Способность к регенерации позволяет животным отрастить новый хвост. То есть, обе части выживут и на месте среза сформируют хвосты. У передней части в итоге получатся голова и хвост, а у задней – два хвоста.
Но при условии, что с каждой стороны осталось достаточное число сегментов. Способность к регенерации позволяет животным отрастить новый хвост. То есть, обе части выживут и на месте среза сформируют хвосты. У передней части в итоге получатся голова и хвост, а у задней – два хвоста.
Голову вырастить хвостовая половина не способна. Поэтому из двух частей, в лучшем случае, получится один новый червь (второй погибнет от голода).
Размножение дождевого червя делением (вегетативное) – самый распространенный миф о беспозвоночных.
Половой методОрганизм каждой особи одновременно оснащен и мужскими половыми органами (семенниками), и женскими (яичниками). Но без пары черви потомства не дадут, так как от природы наделены перекрестным синхронным гермафродитизмом.
Размножение дождевого червя поэтапно:
- С 3-4 месяцев жизни, иногда раньше, на теле червей появляется светлое утолщение в области 32-37 сегментов, называемое пояском. Это признак, что животное созрело для размножения.
- Готовый к спариванию червь находит партнера для взаимного оплодотворения. Особи соединяются брюшками для обмена половыми клетками. В пояске осуществляется построение кокона, внутри которого из собственных яйцеклеток за пару дней созревают яйца.
- Готовые яйца червь откладывает в муфту – слизистое образование поверх тела. Туда же поступает полученная от партнера семенная жидкость.
- Червь снимает через голову загустевшую муфту и оставляет в земле, где оплодотворение завершается.
- На вторые сутки слизистая муфта превращается в прочный кокон, в котором 2-3 недели развиваются оплодотворенные яйца, а затем зародыши. Если погода холодная, то развитие растягивается даже на 3-5 месяцев, но содержимое кокона надежно защищено от внешних факторов.
- На свет появляются крошечные, но уже готовые к самостоятельной жизни червячки.
Условия для роста и размноженияВ хороших условиях
каждые 5-7 суток черви спариваются и откладывают новые коконы
Активная жизнедеятельность и размножение червей начинаются, когда почва прогревается до +8°С. В природе во время холодов и засухи животные уходят вглубь почвы и впадают в анабиоз до наступления благоприятных времен.
В природе во время холодов и засухи животные уходят вглубь почвы и впадают в анабиоз до наступления благоприятных времен.
Соответственно, для круглогодичного прироста поголовья на вермиферме в первую очередь обеспечивается плюсовая температура, желательно в диапазоне 15-25°С.
Процесс размножения дождевого червя – условия:
- своевременное и грамотное кормление;
- влажность 70-85%;
- кислотность – 6,5-7,5 рН единиц.
Чем благоприятнее и комфортнее условия содержания у питомцев, тем чаще черви приносят потомство.
Иные заводчики считают, что нет разницы между дикими навозниками и селекционными вермикультурами. Однако искусственные виды терпимее к составам кормов, живучее и плодовитее. Беспозвоночные проходят жесткий отбор в экстремальных условиях на протяжении многих поколений, а затем их потомки становятся основателями новой технологической породы, такие как червь старатель.
Для получения биогумуса для домашней рассады подойдут и огородные черви, но для бизнеса или промышленной переработки отходов понадобятся разновидности попродуктивнее.
Методы разведения червейДля вермифермы главное выбрать закрытое пространство, чтобы оградить питомцев от хищных вредителей и вредоносных факторов среды (солнца, дождя, ветра), а заодно и от миграций с места жительства. А непосредственно способ размножения дождевого червя вермифермер выбирает на свой вкус, учитывая дальнейшие планы.
Домашняя мини вермифермаПри желании, червей содержат в квартире или доме, используя любую емкость, вплоть до ведерка из-под майонеза. Маленькая ферма перерабатывает пищевые отходы семьи, а взамен дает удобрение для комнатных растений.
Контейнерный методМобильные и дешевые ящики среднего размера пользуются популярностью в среде вермифермеров. В таких легче переносить питомцев с места на место, собирать биогумус, следить за параметрами.
Если выделить питомцам помещение (подвал, сарай, теплую веранду), то можно компактно разместить хоть сколько червей на маленькой площади, но в 2-4 яруса. Стеллажи громоздки и не отличаются мобильностью, зато дают вероятность расширить производство.
Буртовой метод разведенияПри наличии земельного участка данный способ не ограничивает заводчика в размерах фермы. Разводить червей в буртах выгодно производителям сельскохозяйственной и пищевой продукции, так как собственные отходы производства пойдут на корм питомцам. Кстати, в просторных условиях черви плодятся быстрее, чем в ящиках.
ЗаключениеНеприхотливый земляной червь использует по максимуму возможности, которые предоставляет ему среда обитания, для питания и воспроизведения потомства. Поэтому животные столь распространены в природе и продуктивны в неволе. Приложив минимум усилий, заводчик быстро добьется максимума производительности от питомцев.
Подписывайтесь на нас
Оцените статью
Дождевые черви — архитекторы наших плодородных почв
13. 3. 2020, Lukas Pfiffner
В здоровой почве на одном гектаре луга обитает от одного до трех миллионов дождевых червей. Чем больше червей, тем плодороднее почва. Дождевые черви могут почти полностью исчезнуть из почвы при слишком интенсивном или неправильном ее возделывании. И если разрезать дождевого червя пополам, не получится два червя, как мы часто слышим, выжить может, в лучшем случае, только передняя часть.
Рис. 1: Вылупление червя из кокона
Рис. 2: Взрослый дождевой червь: хорошо различимо железистое образование в виде пояска (clitellum)
Рис. 3: Свернутый кольцом дождевой червь может пережить засуху
За исключением тех червей, которые живут в подстилке на поверхности почвы (особенно компостных дождевых червей), такие поясковые черви (включая обыкновенного дождевого червя) развиваются медленно. Они производят только одно поколение в год, и не более восьми коконов (= яйца дождевого червя). Их продолжительность жизни составляет от двух до восьми лет, в зависимости от вида. Дождевые черви, достигшие половой зрелости, имеют «поясок» — отечные железы, расположенные в передней трети тела, также называемыми клителлюм. Рытье нор и размножение дождевых червей наиболее интенсивны в марте и апреле, а также в сентябре и октябре. Если погода очень сухая и жаркая, дождевые черви засыпают … их летний сон настолько глубокий, что сравним с зимней спячкой ежей. Земляные черви просыпаются осенью, когда погода становится прохладной и влажной. Во время зимних заморозков они уходят вглубь своих коридоров, куда мороз не добирается, и переходят в «режим ожидания». Они возвращаются в активное состояние, когда температура держится на отметке выше нуля в течение нескольких дней. Дождевые черви могут мигрировать на возделываемые поля из нетронутых окружающих почв (например, естественных лугов). Обычный дождевой червь (Lumbricus terrestris) может преодолевать расстояние около двадцати метров в год.
Они производят только одно поколение в год, и не более восьми коконов (= яйца дождевого червя). Их продолжительность жизни составляет от двух до восьми лет, в зависимости от вида. Дождевые черви, достигшие половой зрелости, имеют «поясок» — отечные железы, расположенные в передней трети тела, также называемыми клителлюм. Рытье нор и размножение дождевых червей наиболее интенсивны в марте и апреле, а также в сентябре и октябре. Если погода очень сухая и жаркая, дождевые черви засыпают … их летний сон настолько глубокий, что сравним с зимней спячкой ежей. Земляные черви просыпаются осенью, когда погода становится прохладной и влажной. Во время зимних заморозков они уходят вглубь своих коридоров, куда мороз не добирается, и переходят в «режим ожидания». Они возвращаются в активное состояние, когда температура держится на отметке выше нуля в течение нескольких дней. Дождевые черви могут мигрировать на возделываемые поля из нетронутых окружающих почв (например, естественных лугов). Обычный дождевой червь (Lumbricus terrestris) может преодолевать расстояние около двадцати метров в год.
Дождевые черви могут жить глубоко в почве, в среднем или поверхностном слое почвы
В Швейцарии насчитывается около сорока видов дождевых червей, от четырех до восьми из которых живут в культивируемой почве. Земляные черви предпочитают средне-тяжелые суглинки песчаным суглинкам. Они не любят глину и тяжелые почвы, а также песчаные и сухие почвы, и лишь некоторые из них «специализируются» на кислых торфяных почвах, то есть виды, которые приспособились к таким специфическим враждебным условиям. Различные типы дождевых червей можно разделить примерно на три экологические группы (см. график и таблицу):
Анектические (глубокороющие) виды, то есть те, которые роют вертикальные, глубокие ходы, важны для сельскохозяйственных почв. Их красноватый цвет защищает их от воздействия ультрафиолетовых лучей (рис. 2). Эти дождевые черви роют вертикальные, устойчивые ходы диаметром от 8 до 11 мм, где они остаются при благоприятных условиях на протяжении всей своей жизни.
Эндогейные (подповерхностные) виды, то есть те, которые роют ходы под поверхностью почвы, бледны и прозрачны. Эти дождевые черви, которые роют, в основном, горизонтальные неровные ходы, почти никогда не достигают поверхности земли. Молодых дождевых червей обычно можно найти довольно высоко в корнях растений.
Эпигейные (поверхностные) виды, то есть живущие в поверхностных слоях, относительно редко встречаются на пахотных землях, потому что они не могут жить вне устойчивого слоя почвы.
График 1: Слои почвы, в который встречаются три основных группы дождевых червей.
Таблица 1: Экологические группы дождевых червей
| Анектики | Эндогеики | Эпигеики | |
| Определение | Виды, роющие глубокие вертикальные коридоры | Виды, роющие горизонтальные ходы в подповерхностном слое почвы | Виды, живущие в поверхностном слое почвы |
| Место обитания | Все слои почвы глубже 3-4 м (лессовидный суглинок) | Плодородный слой почвы (5-40 см), гуминовые минеральные почвы | Поверхностная подстилка, особенно в лугах и лесах |
| Размер | Как правило, большого размера, 15-45 см длиной | Небольшого размера или до 18 см длиной | Маленького размера, как правило, 2-6 см длиной |
| Пища | Затаскивают крупные остатки растений в ходы, где они живут | Oстатки растений, смешанные с перегноем | Небольшие кусочки растений с поверхности почвы |
| Размножение | Ограничено | Ограничено | В большом количестве |
| Продолжительность жизни | Длинная: 4 — 8 года | Средняя: 3 — 5 года | Короткая: 1–2 года |
| Чувствительность к свету | Умеренная | Сильная | Легкая |
| Цвет | Красно-коричневый, голова более темная | Светлый | Обычно красно-коричневый |
| Примеры | Обыкновенный дождевой червь, Nicrodilus Géardi | Белый червь (Octolasium lacteum), серый червь (Allolobophora caliginosa) | Компостный червь, красный дождевой червь |
Пища имеет решающее значение для развития дождевых червей, которые в основном питаются растительными остатками. Ночью они пасутся на «пастбище из водорослей», которое образуется в течение дня, и затаскивают растительные остатки в свои ходы, где микроорганизмы «предварительно переваривают» их через две-четыре недели. Дождевые черви не имеют зубов и поэтому не могут жевать корни растений. Например, обыкновенный дождевой червь подхватывает растительные остатки, которые находятся ниже или на поверхности почвы.
Рис. 4: Ходы дождевых червей, покрытые фекалиями (коричневые пятна — это следы гумуса), также содержащих белые точки, которые являются кристаллизованными питательными веществами.
Дождевые черви называются архитекторами, а также инженерами плодородной почвы. Их влияние очень разнообразно. Они откладывают большое количество (ежегодно около 40-100 тонн на гектар) фекалий в почву (приблизительно 40%) и на поверхность почвы (приблизительно 60%).
Дождевые черви вентилируют почву
Ходы, вырытые дождевыми червями, обеспечивают хорошую аэрацию почвы и увеличивают долю макропор.
Дождевые черви улучшают способность почвы впитывать водуПлотные вертикальные коридоры анектических видов (например, обыкновенного дождевого червя) особенно улучшают поглощение, хранение, просачивание и отвод воды в почве, что значительно способствует предотвращению эрозии почвы потоками воды. В непахотных почвах имеется до 150 каналов на м2, что составляет 900 метров ходов на м2 на глубине 1 метра. Эти вертикальные коридоры, укрепленные слизью, выделяемой дождевыми червями, могут достигать глубины до 3 метров в глубоких лессовых почвах и даже до 6 метров в черноземах.
Дождевые черви утилизируют растительные остаткиНа полях до 6 тонн мертвого органического вещества на гектар ежегодно вносится в почву дождевыми червями, а в лесах ежегодно обрабатывается до девяти тонн палых листьев на гектар.
Рис. 5: Дождевые черви откладывают большое количество кучек фекалий, называемых выбросами, на поверхности почвы — дождевые черви активны и практически не приводят к образованию трещин в почве (почва, обрабатываемая в органическом сельском хозяйстве в течение 20 лет в рамках теста Therwil DOC).
Рис. 6: Здесь выбросы практически отсутствуют. Почва имеет тенденцию к растрескиванию (обработанная почва при комплексном возделывании в течение 20 лет в рамках теста Therwil DOC).
Фекалии дождевого червя представляют собой смесь растительных и минеральных частиц; они содержат высококонцентрированные питательные вещества в форме, легко усваиваемой растениями. Ежегодно дождевые черви производят от 40 до 100 тонн фекалий на гектар. Такой очень ценный гумус откладывается на почве в виде выбросов, которые содержат примерно в 5 раз больше азота, в 7 раз больше фосфора и в 11 раз больше калия, чем окружающая почва.
Дождевые черви обновляют почвуЗемляные черви извлекают минеральные вещества из нижнего слоя почвы и транспортируют их в верхний слой почвы, за счет чего он постоянно обновляется.
Дождевые черви «дезинфицируют» почвуДождевые черви способствуют имплантации и размножению бактерий и эдафических (подповерхностных) грибов в своих ходах и выбросах. Как только они удаляют остатки палых листьев в почве, вредные организмы, которые населяют их (зимующие формы патогенных грибов, таких как парша яблока и эпикоккоз, или гусеницы, образующие коридоры в листьях), подвергаются биологическому разложению. Однако устойчивые формы выживают в фекалиях дождевого червя и затем могут обнаруживаться в выбросах.
Дождевые черви способствуют росту корнейВ более, чем девяноста процентов случаев в ходах червей присутствуют корни растений, поскольку ходы позволяют им проникать глубоко в почву и находить идеальные условия для роста (экскременты дождевых червей, богатые питательными веществами, вода) без каких-либо помех.
Земляные черви способствуют созданию и стабильности почвенных агрегатовВырабатывая слизь и энергично перемешивая органические вещества с минеральными частицами почвы и микроорганизмами, дождевые черви не только создают устойчивую комковатую структуру, которая помогает уменьшить трещинообразование в почве и облегчает ее разрыхление, но и способствуют более эффективному удержанию питательных веществ и воды. Таким образом, производство большого количества фекалий разрыхляет тяжелые почвы и улучшает сцепление песчаных почв.
Анектические дождевые черви способны проникать в уплотненные слои почвы и улучшать вертикальный поток воды.
Рис. 7: Вспашка борозды помогает избежать уплотнения почвы под верхним слоем.
Рис. 8: Отказ от использования комбинаций тяжелых машин помогает защитить дождевых червей.
Разумный подход к вспахиванию почвы
- Используйте плуги и ротационные бороны любых типов только в случае крайней необходимости. В зависимости от периода, их использование приводит к массовому уничтожению популяции дождевых червей, до 25 процентов после вспашки и до 70 процентов после прохождения ротационной бороны.
- Избегайте интенсивной обработки почвы во время основных периодов активности дождевых червей (март-апрель и сентябрь-октябрь).
- Обработка сухих или холодных почв наносит гораздо меньше вреда дождевым червям, так как большинство из них при таких условиях находятся в глубоких слоях почвы.
- Следует перемешивать почву как можно меньше. Однако, если вспашка необходима, она должна быть ограничена поверхностной вспашкой, что позволит избежать уплотнения глубоких слоев почвы.
- Выбирайте наиболее щадящие методы обработки почвы и комбинации сельскохозяйственных машин.
- Обработка почвы допускается только в том случае, если она достаточно сухая и имеет хорошую несущую способность.
- Продумайте степень использования механизмов так, чтобы минимизировать давление на почву. Чем тяжелее машины, тем сильнее происходит уплотнение почвы, что отрицательно влияет на количество дождевых червей и других живых микроорганизмов.
- Обильное поступление разнообразных растительных остатков в почву является залогом богатства биологического спектра почвы. Этому способствуют разнообразие растительных остатков и севооборот различных культур с постоянным внесением сидератных удобрений. Если дождевых червей хорошо кормить, их популяция не только выживет, но и будет расти.
- Почва всегда должна быть покрыта растительными остатками.
- Растительный покров почвы, особенно во время зимовки, чрезвычайно полезен для дождевых червей и всей локальной почвенной фауны.
- Многолетние луга и луга с бобовым травостоем восстанавливают популяции дождевых червей и являются более полезными в этом плане, чем однолетние луга.
Разбавленный или подготовленный жидкий навоз, используемый в умеренных количествах и в нужное время, также способствует созданию благоприятных условий для дождевых червей.
Тип и количество органических удобрений влияющие на дождевых червей:
- Хорошо удобренная сбалансированная почва благоприятна для растений и дождевых червей. Свежий компост более благоприятен, чем выдержанный компост, потому что выдержанный компост содержит меньше пищи для дождевых червей.
Органические удобрения должны вноситься под поверхность почвы.
Жидкий навоз: в разбавленном и / или предварительно подготовленном виде оказывает положительное влияние на популяцию дождевых червей. Аммиак, содержащийся в неподготовленном жидком навозе, особенно когда почва насыщена водой, может серьезно повредить дождевым червям, которые живут и / или находятся на поверхности почвы.
Жидкий навоз следует вносить только в почву, способную его принять.
Умеренное внесение (приблизительно 25 м3 / га) жидкого навоза благотворно влияет на дождевых червей.
Периодически необходимо проводить известкование, руководствуясь результатами измерений уровня pH.
- Растительные остатки, застрявшие глубоко в почве
- Плохо вентилируемая, уплотненная и затопленная почва
- Кислая почва с уровнем pH ниже 5.5
Популяция от 120 до 140 дождевых червей на квадратный метр представляет собой оптимальную плотность для возделываемых почв Швейцарского плато. Вы можете приблизительно оценить количество дождевых червей в почве следующими способами
Выкапывание:При выкапывании участка почвы размером 10 x 10 см на глубину 25 см в средне-тяжелой плодородной суглинистой почве должно содержаться от одного до двух дождевых червей (что соответствует 100–200 дождевым червям на квадратный метр).
Подсчет выбросов:Выбросы подсчитываются на площади 50 х 50 см во время основных периодов активности дождевых червей (март-апрель и сентябрь-октябрь):
- До 5: Низкая активность дождевых червей, почва содержит небольшое количество дождевых червей.
- 10: Средняя активность дождевых червей.
- 20 и более: Высокая активность дождевых червей, почва содержит большое количество дождевых червей.
Программа вступительных испытаний по биологии
Программа составлена на основе федерального государственного образовательного стандарта среднего общего образования и объединяет в себе все основные содержательные компоненты биологических знаний. В содержание программы включен материал из всех разделов школьной биологии. Экзаменационные задания по биологии не выходят за рамки данной программы, но требуют глубокой проработки всех ее элементов. Для успешных ответов на задания необходимо свободное и осознанное владение биологическими понятиями, теориями, законами и закономерностями. В содержание вступительных испытаний включены задания в тестовой форме.
ПОРЯДОК ПРОВЕДЕНИЯ ВСТУПИТЕЛЬНЫХ ИСПЫТАНИЙ
В определенное расписанием время абитуриенты должны занять места в назначенной аудитории, для чего с собой необходимо иметь: паспорт, экзаменационный лист, ручку. После размещения всех, допущенных к вступительным испытаниям, представитель экзаменационной комиссии объясняет правила оформления ответа и раздает листы с экзаменационными заданиями. С этого момента начинается отсчет времени. Продолжительность вступительных испытаний — два академических часа (90 минут).
ТРЕБОВАНИЯ К ПИСЬМЕННОМУ ЭКЗАМЕНУ И КРИТЕРИИ ОЦЕНКИ
Абитуриент, сдающий вступительный экзамен в вуз по биологии должен показать знания, навыки и умения, соответствующие программе средней общеобразовательной школы.
Письменный экзамен по биологии состоит из 20-ти тестовых заданий.
Правильное решение каждого задания оценивается 5 баллами.
Результаты выполнения экзаменационной работы оцениваются по 100-балльной системе. Максимальное количество баллов, выставляемых за экзаменационную работу, – 100. Количество баллов, необходимое для получения положительной оценки, — 36.
СОДЕРЖАНИЕ ПРОГРАММЫ
Растения. Бактерии. Грибы. Лишайники. Растительный мир как составная часть природы, его разнообразие. Значение растений в природе и жизни человека. Охрана растений. Корень. Развитие корня из семени. Виды корней. Типы корневых систем. Внешнее и внутреннее строение корня. Рост корня. Дыхание корней. Основные функции корня. Удобрения. Значение обработки почвы, внесение удобрений и полив. Видоизменения корней.
Побег. Почка – зачаточный побег. Почки листовые и цветочные. Строение почек. Развитие побега из почки. Лист. Внешнее строение листа. Жилкование. Листья простые и сложные. Листорасположение. Особенности микроскопического строения листа в связи с его функциями. Фотосинтез. Дыхание листа. Испарение воды листьями. Видоизменения листьев.
Стебель. Морфологические формы стеблей. Ветвление и формирование кроны. Внутреннее строение стебля древесного растения в связи с выполняемыми функциями. Рост стебля в длину и в толщину. Передвижение минеральных и органических веществ по стеблю. Видоизмененные побеги: корневище, клубень, луковица; их биологическое значение.
Способы вегетативного размножения цветковых растений. Вегетативное размножение с помощью корней, листьев, стеблей и видоизмененных побегов. Значение вегетативного размножения.
Цветок – орган семенного размножения. Строение цветка: цветоножка, цветоложе, околоцветник, тычинки и пестик. Однополые и обоеполые цветки. Однодомные и двудомные растения. Значение цветка в размножении растений. Соцветия, их многообразие и биологическое значение. Перекрестное опыление, самоопыление. Ветроопыляемые и насекомоопыляемые растения. Образование плодов и семян. Типы плодов. Строение семян однодольных и двудольных растений. Химический состав семян. Дыхание семян. Питание и рост проростков. Распространение плодов и семян.
Растение – целостный организм. Ткани растительного организма. Взаимосвязи клеток, тканей и органов. Основные процессы жизнедеятельности растения. Растительные сообщества.
Основные отделы растений.
Водоросли. Строение и жизнедеятельность одноклеточных и нитчатых зеленых водорослей. Размножение водорослей. Бурые морские водоросли. Значение водорослей в природе и жизни человека.
Мхи. Строение и размножение (на конкретном примере), усложнение в процессе эволюции. Особенности строения сфагнума. Образование торфа и его значение.
Папоротникообразные. Строение и размножение (на конкретном примере), усложнение в процессе эволюции. Папоротники, хвощи, плауны. Роль в природе.
Голосеменные. Строение и размножение (на конкретном примере), усложнение в процессе эволюции. Многообразие. Значение в природе и в хозяйственной деятельности человека.
Покрытосеменные. Особенности строения и жизнедеятельности покрытосеменных, их классификация. Признаки классов двудольных и однодольных растений. Класс Двудольные растения. Характеристика семейств крестоцветных, розоцветных, бобовых, пасленовых и сложноцветных, их значение в природе и жизни человека. Класс Однодольные растения. Семейства: лилейные, злаки. Отличительные признаки растений данных семейств, их биологические особенности и значение.
Происхождение культурных растений. Понятие о сорте. Важнейшие сельскохозяйственные растения (зерновые, плодово-ягодные, овощные, масличные, технические и другие). Биологические основы их выращивания.
Бактерии. Особенности строения и жизнедеятельности прокариотической клетки. Формы бактерий. Размножение бактерий. Распространение бактерий. Роль бактерий в природе и жизни человека.
Общая характеристика царства грибов. Особенности строения и жизнедеятельности шляпочных грибов. Дрожжи. Плесневые грибы: пеницилл и мукор. Грибы-паразиты. Их строение, питание и размножение. Значение грибов в природе и хозяйственной деятельности человека.
Лишайники. Особенности строения, питания и размножения лишайников как симбиотических организмов. Многообразие лишайников. Роль лишайников в природе и жизни человека.
Животные. Основные отличия животных от растений. Черты их сходства. Многообразие животного мира.
Одноклеточные. Общая характеристика. Среда обитания, особенности строения и жизнедеятельности одноклеточных животных: передвижение, питание, дыхание, размножение (на примере амебы обыкновенной, инфузории-туфельки). Образование цист. Многообразие и значение одноклеточных животных.
Тип Кишечнополостные. Общая характеристика типа. Гидра пресноводная: среда обитания, внешнее строение, вид симметрии. Дифференциация клеток у кишечнополостных. Питание, передвижение, регенерация и размножение. Многообразие кишечнополостных (коралловые полипы и медузы), их значение.
Тип Плоские черви. Класс Ресничные черви. Белая планария: среда обитания, внешнее строение, передвижение. Двусторонняя симметрия. Ткани, органы, системы органов плоских червей. Классы Сосальщики и Ленточные черви. Особенности строения и жизнедеятельности в связи с паразитическим образом жизни. Жизненные циклы печеночного сосальщика и бычьего цепня. Многообразие плоских червей. Вред, наносимый животноводству и людям, меры борьбы.
Тип Круглые черви. Аскарида человеческая: внешнее и внутреннее строение, жизнедеятельность и размножение. Профилактика аскаридоза.
Тип Кольчатые черви. Общая характеристика типа. Малощетинковые черви. Дождевой червь: среда обитания, внешнее и внутреннее строение. Регенерация, размножение. Роль дождевых червей в почвообразовании. Многощетинковые черви. Усложнение кольчатых червей в процессе эволюции.
Тип Моллюски. Общая характеристика типа. Среда обитания. Особенности строения, питания, дыхания, размножения на примере одного из представителей типа. Многообразие моллюсков: классы Брюхоногие, Двустворчатые и Головоногие. Значение в природе и жизни человека.
Тип Членистоногие. Общая характеристика типа Членистоногие.
Класс Ракообразные. Общая характеристика класса. Среда обитания, особенности строения, жизнедеятельности, размножения. Многообразие. Роль в природе и жизни человека.
Класс Паукообразные. Общая характеристика класса. Особенности строения, питания, дыхания, размножения и поведения паука-крестовика в связи с жизнью на суше. Клещи. Особенности строения и жизнедеятельности. Роль в природе и жизни человека. Меры защиты от клещей.
Класс Насекомые. Общая характеристика класса. Особенности строения, и жизнедеятельности (на конкретном примере). Размножение. Типы развития насекомых.
Характеристика основных отрядов насекомых: Чешуекрылых, Двукрылых, Перепончатокрылых. Значение насекомых.
Тип Хордовые. Общая характеристика и происхождение хордовых. Особенности строения ланцетника. Черты сходства с беспозвоночными животными.
Хрящевые и костные рыбы. Общая характеристика класса. Особенности внешнего и внутреннего строения в связи со средой обитания. Нервная система и органы чувств. Рефлексы. Поведение. Размножение и развитие. Древние кистеперые рыбы – предки Земноводных. Многообразие и значение рыб.
Класс Земноводные. Общая характеристика класса. Многообразие земноводных (отряды хвостатые, бесхвостые), их происхождение, значение и охрана. Особенности строения, жизнедеятельности и размножения в связи с обитанием в воде и на суше. Многообразие и значение земноводных.
Класс Пресмыкающиеся. Общая характеристика класса. Среда обитания, особенности строения, размножения, поведения в связи с жизнью на суше. Регенерация. Происхождение пресмыкающихся. Древние пресмыкающиеся. Многообразие пресмыкающихся, их значение и охрана.
Класс Птицы. Общая характеристика класса. Внешнее строение. Особенности внутреннего строения и процессов жизнедеятельности, связанные с полетом. Поведение птиц. Размножение и развитие, забота о потомстве. Приспособленность птиц к сезонным явлениям в природе. Экологические группы птиц. Происхождение птиц. Роль птиц в природе и жизни человека. Охрана птиц.
Класс Млекопитающие. Общая характеристика класса. Особенности внешнего строения, скелета, мускулатуры, внутреннего строения и обмена веществ. Размножение, развитие, забота о потомстве. Приспособленность млекопитающих к сезонным явлениям в природе. Усложнение нервной системы, органов чувств, поведения. Происхождение млекопитающих. Первозвери. Сумчатые. Характеристика отрядов плацентарных (насекомоядные, рукокрылые, грызуны, зайцеобразные, хищные, ластоногие, китообразные, копытные, приматы). Роль млекопитающих в природе и жизни человека, их охрана.
Человек и его здоровье. Особенности строения и жизнедеятельности животной клетки. Основные ткани организма человека. Органы и системы органов, связь их строения с выполняемыми функциями. Нервная и гуморальная регуляция функций физиологических систем. Человек и окружающая среда. Опорно-двигательная система человека. Сходство скелета человека и животных. Отделы скелета человека. Особенности скелета человека, связанные с трудовой деятельностью и прямохождением. Состав и строение костей. Рост костей. Типы соединения костей и их значение. Первая помощь при ушибах, растяжениях связок, вывихах и переломах. Мышцы, их строение и функции. Основные группы мышц. Работа мышц. Влияние ритма и нагрузки на работу мышц. Утомление мышц. Предупреждение искривления позвоночника и развития плоскостопия. Кровь и кровообращение. Внутренняя среда организма и ее относительное постоянство. Значение крови и кровообращения. Состав крови, плазма крови. Свертывание крови. Строение и функции клеток крови. Группы крови, переливание крови, донорство. Иммунитет. Значение прививок. Органы кровообращения: сердце и кровеносные сосуды. Строение и работа сердца. Большой и малый круги кровообращения. Движение крови по сосудам, кровяное давление, пульс. Регуляция деятельности сердца и кровеносных сосудов. Лимфообращение. Предупреждение сердечно-сосудистых заболеваний. Первая помощь при кровотечениях. Вредное влияние курения на сердечно-сосудистую систему. Дыхательная система. Значение, строение и функции органов дыхания. Голосовой аппарат. Дыхательные движения. Газообмен в легких и тканях. Жизненная емкость легких. Нервно-гуморальная регуляция дыхания. Искусственное дыхание. Инфекционные болезни, передающиеся через воздух. Предупреждение воздушно-капельных инфекций. Гигиена дыхания. Влияние загрязнения воздушной среды на дыхательную систему человека.
Пищеварительная система. Питательные вещества и пищевые продукты. Понятие о пищеварении. Роль ферментов в пищеварении. Строение и функции органов пищеварения. Пищеварение в ротовой полости, желудке и кишечнике. Всасывание. Значение печени и поджелудочной железы в пищеварении. Регуляция процессов пищеварения. Роль И.П. Павлова в изучении функций органов пищеварения. Гигиена питания.
Общая характеристика обмена веществ и энергии. Пластический и энергетический обмены, их взаимосвязь. Обмен белков, жиров, углеводов в организме человека. Водно-солевой обмен. Витамины, их значение в обмене веществ. Авитаминозы, гиповитаминозы, гипервитаминозы. Рацион, режим и нормы питания.
Мочевыделительная система: строение и функции. Образование мочи. Значение выделения продуктов обмена. Профилактика заболеваний органов мочевыделения.
Строение и функции кожи. Роль кожи в терморегуляции. Закаливание организма. Гигиена кожи. Первая помощь при тепловом и солнечном ударах, ожогах и обморожениях.
Железы внутренней секреции. Их значение в жизнедеятельности и развитии организма. Гормоны. Внутрисекреторная деятельность гипофиза, щитовидной железы, надпочечников, поджелудочной железы. Болезни, связанные с нарушением деятельности желез внутренней секреции.
Нервная система. Ее значение в регуляции и координации функций организма и осуществлении взаимосвязи организма со средой. Центральная и периферическая нервная система человека. Понятие о рефлексе. Строение и функции спинного мозга. Строение и функции отделов головного мозга. Большие полушария головного мозга: доли и функциональные зоны. Роль вегетативной нервной системы в регуляции работы внутренних органов. Вредное влияние никотина, алкоголя и наркотиков на нервную систему. Анализаторы, органы чувств, их значение. Строение, функции и гигиена зрительного и слухового анализаторов. Высшая нервная деятельность человека (ВНД). Роль И.М. Сеченова и И.П. Павлова в создании учения о высшей нервной деятельности. Безусловные и условные рефлексы. Биологическое значение образования и торможения условных рефлексов. Отличия высшей нервной деятельности человека от ВНД животных. Речь и мышление. Сознание как функция мозга. Социальная обусловленность поведения человека. Сон, его значение и гигиена умственного труда. Режим дня и его значение.
Система органов размножения: строение и функции. Оплодотворение и внутриутробное развитие. Рождение ребенка. Уход за новорожденным. Личная гигиена подростков.
Общая биология. Биология – наука о жизни. Методы биологических исследований. Уровни организации живой материи. Основы цитологии. Клетка – структурная и функциональная единица живого. Химические элементы клеток. Неорганические соединения клетки. Роль воды в клетке и организме. Органические вещества клеток: углеводы, липиды, белки, нуклеиновые кислоты, АТФ; их роль в клетке.
Строение эукариотической клетки. Клеточная стенка и наружная клеточная мембрана: строение и функции. Протоплазма и цитоплазма клетки. Ядро: строение и функции. Хромосомы и хроматин. Вакуолярная система клетки: эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли). Митохондрии и пластиды. Немембранные органоиды: рибосомы, цитоскелет, клеточный центр, органоиды движения. Клеточные включения. Особенности строения клеток прокариот. Сравнительная характеристика строения клеток растений и животных. Особенности строения и жизнедеятельности прокариот. Положения клеточной теории.
Обмен веществ и превращение энергии – основа жизнедеятельности клетки. Классификация организмов по источнику энергии и типу питания. Энергетический обмен в клетке (на примере окисления глюкозы), его сущность, значение АТФ. Фотосинтез: световая и темновая фазы, значение. Хемосинтез. Ген, генетический код и его свойства. Биосинтез белков: транскрипция и трансляция. Реакции матричного синтеза. Взаимосвязь пластического и энергетического обменов.
Неклеточные формы жизни (вирусы). Строение, размножение вирусов. Вирусные заболевания человека. Профилактика СПИДа.
Размножение и индивидуальное развитие организмов. Соматические и половые клетки многоклеточного организма. Хромосомы, гаплоидный и диплоидный набор хромосом. Гомологичные хромосомы.
Клеточный цикл. Интерфаза. Механизм и биологическое значение митоза. Мейоз – редукционное деление, механизм и биологическое значение мейоза.
Сперматогенез и овогенез у животных. Оплодотворение у животных. Двойное оплодотворение цветковых растений. Онтогенез. Эмбриональное и постэмбриональное развитие. Прямое и непрямое постэмбриональное развитие. Формы размножения организмов: бесполое и половое. Способы бесполого размножения (митоз, спорообразование, фрагментация, почкование, вегетативное размножение). Половое размножение организмов. Партеногенез. Гермафродитизм.
Основы генетики. Генетика как наука, ее задачи и методы исследования. Основные понятия генетики: наследственность, изменчивость, доминантные и рецессивные признаки. Аллельные гены. Фенотип и генотип. Гетерозиготные и гомозиготные организмы. Закономерности, установленные Г. Менделем для моно – и дигибридного скрещиваний: правило единообразия гибридов, закон расщепления, гипотеза чистоты гамет, закон независимого наследования признаков. Их цитологические основы. Генотип как целостная исторически сложившаяся система. Взаимодействие неаллельных генов. Анализирующее скрещивание. Сцепленное наследование. Хромосомная теория наследственности. Генетика пола. Наследование признаков, сцепленных с полом. Генетика человека. Методы изучения наследственности человека. Наследственные болезни человека. Значение генетики для медицины и здравоохранения.
Наследственная и ненаследственная изменчивость. Классификация мутаций. Мутагены. Искусственный мутагенез. Мутации – материал для естественного и искусственного отбора. Закон гомологических рядов в наследственной изменчивости Н.И. Вавилова. Модификационная изменчивость. Норма реакции. Статистические закономерности модификационной изменчивости.
Основы селекции. Селекция как наука, ее задачи. Значение работ Н.И. Вавилова для развития селекции. Основные методы селекции растений. Особенности и методы селекции животных. Типы разведения животных. Методы селекции микроорганизмов. Биотехнология, ее значение и основные направления: микробиологический синтез, генная и клеточная инженерия.
Основы экологии. Экология: предмет, задачи и методы исследования. Среда обитания. Особенности водной, наземно-воздушной, почвенной и организменной сред обитания. Приспособленность организмов к обитанию в различных средах. Экологические факторы: абиотические, биотические, антропогенные; их комплексное воздействие на организм. Закономерности влияния экологических факторов на организм.
Экологическая характеристика популяции. Структура популяции. Динамика численности популяции. Факторы, регулирующие численность. Взаимоотношения особей в популяциях.
Понятие о сообществе, биогеоценозе, экосистеме. Структура и организация биогеоценоза. Разнообразие популяций в биогеоценозе, их взаимосвязи. Типы взаимодействий особей в биогеоценозах. Цепи питания и трофические сети. Саморегуляция и устойчивость биогеоценозов. Смена биогеоценозов. Агроценозы. Охрана биогеоценозов.
Основы эволюционного учения. Додарвиновский период в биологии: значение трудов К.Линнея и Ж.-Б. Ламарка. Предпосылки возникновения учения Дарвина. Основные положения теории эволюции Ч. Дарвина.
Популяция – единица вида и эволюции. Движущие силы эволюции: естественный отбор, борьба за существование, наследственная изменчивость, мутации, изоляция, дрейф генов, популяционные волны, генный поток. Естественный отбор – ведущий эволюционный фактор. Форы отбора. Механизм возникновения адаптаций. Относительный характер приспособленности.
Вид: критерии и структура вида. Видообразование. Доказательства эволюции. Биологический прогресс и регресс. Пути достижения биологического прогресса.
Развитие органического мира. Происхождение жизни на Земле. Развитие мира в архейскую, протерозойскую, палеозойскую, мезозойскую и кайнозойскую эры. Основные ароморфозы.
Происхождение человека. Движущие силы антропогенеза: социальные и биологические факторы. Древнейшие, древние, ископаемые люди современного типа. Человеческие расы, их происхождение и единство.
Основы учения о биосфере. Геосферы Земли. Биосфера и ее границы. В.И. Вернадский о возникновении биосферы. Живое, косное и биокосное вещество, их свойства. Функции живого вещества, роль в круговороте веществ и превращении энергии в биосфере. Биосфера в период научно-технического прогресса. Роль человека в природе. Понятие о ноосфере. Загрязнение окружающей среды.
ПОРЯДОК ПРОВЕДЕНИЯ ВСТУПИТЕЛЬНЫХ ИСПЫТАНИЙ
Вступительные испытания по биологии проводятся в письменной форме. В определенное расписанием время абитуриенты должны занять места в назначенной аудитории, для чего с собой необходимо иметь: паспорт, экзаменационный лист, ручку. После размещения всех допущенных к вступительным испытаниям представитель экзаменационной комиссии объясняет правила оформления ответа и раздает листы с экзаменационными заданиями. С этого момента начинается отсчет времени. Продолжительность вступительных 2 академических часа (90 минут). По окончании отведенного времени абитуриенты должны сдать листы ответа представителям экзаменационной комиссии и выйти из аудитории.
После проверки работы выставляется общая балльная оценка, которая сообщается абитуриенту в приемной комиссии.
Правильное решение каждого задания оценивается 5 баллами.
Результаты выполнения экзаменационной работы оцениваются по 100-балльной системе. Максимальное количество баллов, выставляемых за экзаменационную работу – 100. Количество баллов, необходимое для получения положительной оценки, соответствует 45 баллам.
ПРИМЕРЫ ТЕСТОВЫХ ЗАДАНИЙ:
1. Какой уровень организации живого служит основным объектом изучения цитологии?:
1. клеточный
2. популяционно-видовой
3. биогеоценотический
2. Какие формы жизни занимают промежуточное положение между телами живой и неживой природы?:
1. вирусы
2. бактерии
3. лишайники
3. При скрещивании двух морских свинок с черной шерстью (доминантный признак) получено потомство, среди которого особи с белой шерстью составили 25%. Каковы генотипы родителей?:
1. АА х аа
2. Аа х АА
3. Аа х Аа
4. Укажите признак, характерный только для царства растений:
1. имеют клеточное строение
2. дышат, питаются, растут, размножаются
3. имеют фотосинтезирующую ткань
5. Признак приспособленности птиц к полету:
1. появление четырехкамерного сердца
2. образование роговых щитков на ногах
3. наличие полых костей
6. Наибольшее количество крахмала человек потребляет, используя в пищу:
1. листья салата и укропа
2. растительное и сливочное масло
3. хлеб и картофель
7. Совокупность внешних признаков особей относят к критерию вида:
1. географическому
2. морфологическому
3. экологическому
8. У человека в связи с прямохождением:
1. сформировался свод стопы
2. когти превратились в ногти
3. срослись фаланги пальцев стопы
9. Минерализация органических соединений почвы осуществляется благодаря деятельности:
1. наземных животных
2. микроорганизмов
3. корней растений
10. Число хромосом при половом размножении в каждом поколении возрастало бы вдвое, если бы в ходе эволюции не сформировался процесс:
1. митоза
2. оплодотворения
3. мейоза
11. Артериальная кровь превращается у человека в венозную в:
1. капилярах малого круга кровообращения
2. капилярах большого круга кровообращения
3. печеночной вене
12. Организмы в процессе жизнедеятельности постоянно изменяют среду своего обитания, что способствует:
1. росту и развитию организмов
2. саморазвитию экосистемы
3. размножению организмов
13. Какая наука изучает ископаемые остатки организмов:
1. биогеография
2. палеонтология
3. сравнительная анатомия
14. Встраивание своей нуклеиновой кислоты в ДНК клетки-хозяина осуществляют:
1. бактериофаги
2. хемотрофы
3. автотрофы
15. Какой закон проявится в наследовании признаков при скрещивании организмов с генотипами: Аа х Аа?:
1. единообразия
2. расщепления
3. сцепленного наследования
16. Что образуется из оплодотворенной яйцеклетки растений?:
1. семя
2. зародыш
3. эндосперм
17. Млекопитающих можно отличить от других позвоночных по наличию:
1. волосяного покрова и ушных раковин
2. сухой кожи с роговыми чешуями
3. голой кожи, покрытой слизью
18. Колебания уровня сахара в крови и моче человека свидетельствуют о нарушениях деятельности:
1. желудка
2. поджелудочной железы
3. щитовидной железы
19. Интенсивность размножения и ограниченность ресурсов для жизни организмов являются предпосылкой:
1. борьбы за существование
2. мутационной изменчивости
3. изоляции популяций
20. Какой из примеров относят к биотическим факторам?:
1. поедание тлей божьими коровками
2. весенний разлив реки
3. сезонное пересыхание водоема
СПИСОК ЛИТЕРАТУРЫ ДЛЯ ПОДГОТОВКИ К ВСТУПИТЕЛЬНЫМ ИСПЫТАНИЯМ ПО БИОЛОГИИ
1. Билич Г. Л. Биология для поступающих в ВУЗы. – М.: Оникс, 2007.
2. Лемеза Н. Биология для поступающих в ВУЗы. – М.: Юнипресс, 2006.
3. Мамонтов С. Г. Биология. Пособие для поступающих в ВУЗы. – М.: Дрофа, 2001.
4. Чепурнова Н.Е. Биология. Руководство для поступающих в вузы.: Учебное пособие /
5. Шустанова Т.А. Репетитор по биологии для поступающих в ВУЗы. –М.: Феникс, 2008.
Размножение дождевого червя | HowStuffWorks
Одним из самых интересных аспектов дождевых червей является их сексуальность. Дождевые черви — это одновременных гермафродитов , что означает, что черви имеют как мужские, так и женские репродуктивные органы. Во время полового акта среди дождевых червей оба набора половых органов используются обоими червями. Если все идет хорошо, яйца обоих полов оплодотворяются. Вы можете себе представить, что это очень эффективный способ обеспечить выживание вида. Vermicomposters , люди, которые разводят червей и другие организмы в компост, сообщают, что их популяция дождевых червей обычно удваивается каждые 60–90 дней [источник: Вернер].
Для совокупления два червя выстраиваются в линию друг против друга в противоположных направлениях. В этом положении оба червя выделяют столько слизи, что вокруг их тел образуется слизистая трубка . Каждый червь извергает сперму из своих половых органов в эту слизистую трубку, а затем она откладывается в приемнике для спермы другого червя. Акт спаривания завершен, но процесс воспроизводства все еще продолжается, поскольку каждый червь идет своим путем [источник: Конрад].
Вы знаете широкую полосу на передней части любого дождевого червя? Эта полоса называется clitellum , и она отвечает за образование еще одной трубки слизи.Эта полоса продвигается вперед к ротовому концу червя. По мере продвижения вперед слизь проходит по мешочкам, содержащим собственные яйца червя, которые прилипают к слизи. Присоединенные к слизистой трубке, яйца затем проходят через семяприемник, где хранится сперма другого червя. Яйца и сперма соприкасаются в слизистой трубке, и, если все идет хорошо, яйца оплодотворяются.
Полоса слизи отрывается от головы червя и образует кокон в форме лимона для от четырех до 20 яиц червя, которые обычно откладывает обыкновенный европейский дождевой червь.Примерно через две-три недели новорожденные черви вылупятся и вылезут из кокона в почву. Этот цикл воспроизводства может происходить каждую неделю до 10 дней, что еще одна причина, по которой популяции дождевых червей могут расти так быстро [источник: Barrett].
Дождевые черви при необходимости также могут воспроизводить себя. Они могут регенерировать новые сегменты, если потеряют несколько. Большинство дождевых червей регенерируют хвосты лучше, чем головы, но некоторые могут. Однако они не размножаются бесполым путем; только половина (и, вероятно, половина головы) дождевого червя, разделенного на две части, снова превратится в полноценного червя [источник: Томлин].
Размножение компостного червя: как размножаются черви
В системах здорового компостирования обитают одни из самых плодовитых заводчиков: Red Wiggler. Небольшое знание того, как они спариваются и каковы идеальные условия для спаривания, поможет вам собрать больше компоста.
Знаете ли вы, что при благоприятных условиях зрелые красные черви могут производить от двух до трех коконов в неделю? Это много червей, учитывая, что каждый кокон дает от 3 до 20 молодых червей! Если вам нужна долговременная устойчивость, важно, чтобы ваши красные вигглеры были удобными и счастливыми, чтобы стимулировать большее размножение.
Давайте рассмотрим, как размножаются черви и что они могут сделать для вашей системы вермикомпостирования.
Как воспроизводятся красные вигглеры
Большинство людей уже знают, что черви — гермафродиты. Это означает, что у них есть как мужские, так и женские репродуктивные органы. Однако они не могут воспроизводиться в одиночку. Для успешного размножения они должны соединиться с другим червем.
Когда два червя спариваются, они соединяются и начинают выделять слизь, образуя вокруг себя слизистое кольцо.Через несколько часов черви отделяются, откладывают яйца и удобряют внутреннюю часть кокона. Черви выходят из кокона, и он закрывается, оставляя кокон овальной формы, который со временем затвердевает, чтобы защитить детенышей внутри. Через 11 недель птенцы выходят из кокона!
Черви не могут воспроизводиться в любое время. Их тела претерпевают изменения по мере взросления, и примерно в 90 дней они становятся достаточно взрослыми, чтобы спариваться.
Когда красный червь достигает зрелости, на его теле появляется луковичная железа.Вы, наверное, видели это на дождевом черве или ночном ползуне. Эта железа играет важную роль в процессе спаривания, поскольку она производит слизь, необходимую для производства коконов.
Следует обратить внимание на одну важную вещь: чем крупнее черви, тем чаще они спариваются и тем больше вылупившихся птенцов из их коконов. Правильная диета поможет вашим червям вырасти здоровыми и сильными.
Одна из самых приятных частей вермикомпостирования — это самоподдерживающийся элемент.Поскольку черви так много размножаются, у вас никогда не закончится червя в здоровой окружающей среде. Нередко кто-то покупает первую партию красных вигглеров и больше никогда не покупает червей для компостирования.
Существуют противоречивые мнения относительно продолжительности жизни червей, но можно ожидать, что червь сможет использовать как минимум 5 лет, прежде чем они перестанут воспроизводиться. Известно, что дождевые черви живут 10-15 лет, прежде чем умрут, поэтому мы всегда предлагаем помочь червю вернуться во влажную почву, когда вы видите, что он застрял на дорожке, дороге, подъездной дорожке или в другой опасной среде!
Не вдаваясь в подробности, мы надеемся, что это было для вас информативным и поможет собрать более ценный компост из большего количества червей! Просто помните, чем здоровее ваши черви, тем больше они будут размножаться.И чем больше у вас червей, тем больше отливок вы соберете!
Червячная ферма дяди Джима — международная компания по вермикомпостированию, специализирующаяся на красных вигглерах и продуктах для компостирования червей. Это изображение не является нашим собственным.(PDF) Размножение дождевых червей: половой отбор и партеногенез
Casellato S (1987) О полиплоидии олигохатов с особым упором на люмбрицид. In:
Bonvicini AM, Omodeo P (eds) О дождевых червях. Mucchi, Modena, pp 75–87
Casellato S, Rodighiero R (1972) Кариология люмбрицид.3-й вклад. Caryologia
25: 513–524
Cobolli Sbordoni M, De Matthaeis E, Omodeo P, Bidoli R, and Rodino E (1987) Allozyme
изменчивость и дивергенция между диплоидными и триплоидными популяциями Allolobophora 9000eta2: ). В: Bonvicini Pagliai AM, Omodeo P (eds) On Earthworms, pp.
53–74. Избранные симпозиумы и монографии UZI, vol 2. Mucchi, Modena
Delmotte F, Sabater-Mun
~
oz B, Prunier-Leterme N, Latorre A, Sunnucks P, Rispe C, Simon JC
(2003) Философия доказательства гибридного происхождения бесполых линий у видов тлей.
Evolution 57: 1291–1303
Domı
´
nguez J, Velando A, Aira M, Monroy F (2003) Однородительное воспроизведение Eisenia fetida
и E. andrei (Oligochaeta: Lumbrichaeta) осеменение. Pedobiologia
47: 530–534
Edwards CA, Bohlen PJ (1996) Биология и экология дождевых червей, 3-е изд. Chapman and Hall,
London, p 426
Ferna
´
ndez R, Novo M, Gutie
´
rrez M, Almodo
´
var A, Dı 9´20007 9000
´
n D (2010) Жизненный цикл и воспроизводимость —
признаков дождевого червя Aporrectodea trapezoides (Duge
`
s, 1828) в лабораторных культурах.
Pedobiologia 53: 295–299
Field SG, Michiels NK (2005) Паразитизм и рост дождевого червя Lumbricus terrestris:
Затраты на приспособляемость грегаринового паразита Monocystis sp. Parasitology 130: 1–7
Field SG, Schirp HJ, Michiels NK (2003) Влияние инфекции Monocystis sp на рост и
брачное поведение дождевого червя Lumbricus terrestris. Can J Zool 81: 1161–1167
Garvı
´
n MH, Trigo D, Herna
´
ndez P, Dı
´
az Cosı
´
(2003) и размножениеHormogaster elisae (Oligochaeta, Hormogastridae).Inv Biol 122: 152–157
Gates GE (1972) Вклад в распространение дождевых червей в Северной Америке. № 3. IV. Виды трапеции
группа. Bull Tall Timbers Res Stn 12: 146
Gates GE (1974) Вклады в распространение североамериканских дождевых червей. № 10. Вклад в пересмотр
Lumbricidae X: Dendrobaena octaedra (Savigny) 1826 со специальной ссылкой
на важность его партеногенетического полиморфизма для классификации
дождевых червей.Bull Tall Timbers Res Stn 15:57
Grigoropoulou N, Butt KR, Lowe CN (2008) Воздействие на взрослых особей Lumbricus terrestris на коконы и
детенышейв ящиках Эванса. Pedobiologia 51: 343–349
Grove AJ (1925) О репродуктивных процессах дождевого червя Lumbricus terrestris.QJ
Microsc Sci 69: 245–290
Grove AJ, Cowley LF (1926) О репродуктивных процессах червяк-бренди, Eisenia fetida
(сб.). Q J Microsc Sci 70: 559–581
Haimi J, Knott KE, Yliniemi I (2007) Влияет ли металлическое загрязнение на клональное разнообразие партеногенетического земляного червя
Dendrobaena octaedra? Eur J Soil Boil 43: 268–272
Hale CM, Frelich LE, Reich PB (2005) Динамика инвазии экзотических европейских дождевых червей в северных
лиственных лесах штата Миннесота, США.Ecol Appl 15: 848–860
Heethoff M, Etzold K, Scheu S (2004) Митохондриальные последовательности COII указывают на то, что партено-
генетический дождевой червь Octolasion tyrtaeum (Savigny 1826) состоит из двух линий, различающихся размером тела
и генотипом. . Pedobiologia 48: 9–13
Hughes RN (1989) Функциональная биология клональных животных. Chapman and Hall, London, p. 330
ICZN (1999) Международный кодекс зоологической номенклатуры (4-е издание). Международный фонд
Зоологическая номенклатура, c / o Музей естественной истории, Лондон, стр. 306
Янике Дж., Селандер Р.К. (1979) Эволюция и экология партеногенеза у дождевых червей.Am
Zool 19: 729–737
Jaenicke J, Parker ED, Selander RK (1980) Структура клональной ниши в партеногенетической земле —
червь Octolasion tyrtaeum
. Am Nat 116: 196–205
Jamieson BGM (1992) Oligochaeta. В: Harrison FW, Gardiner SL (eds) Miscroscopic Anatomy
of Invertebrates, vol 7. Wiley-Liss, New York
84 D.J. Dı
´
az Cosı
´
n et al.
Репродуктивная система дождевого червя — Онлайн-биологические заметки
7 ноября 2020 Гаураб Карки Зоология 0
Репродуктивная система дождевого червяРазмножение дождевого червя
- Дождевые черви бывают однодомными, гермафродитными или бисексуальными.
- Однако самооплодотворение не происходит из-за относительного расположения мужских и женских репродуктивных органов, а также из-за протандральности.
- Протандрус относится к состоянию, при котором мужские репродуктивные органы созревают раньше, чем женские.
- Таким образом, у дождевых червей происходит перекрестное оплодотворение.
- Он состоит из мужских репродуктивных органов и женских репродуктивных органов.
- Мужские репродуктивные органы состоят из семенников, семенников, семенных пузырьков, семявыносящей воронки, семявыносящего протока, предстательной железы и добавочных желез.
- Мужские репродуктивные органы включают в себя:
| Структура | Номер | Расположение | ||||||||||||||||||
| Testis | 2 пары | |||||||||||||||||||
| Яичек | 1 пара | 10 -й и 11 -й сегмент | ||||||||||||||||||
| Семенные пузырьки | 2 пары | 11 -й и 12 -й сегмент 9024elsm | сегмент 2 пары10 th и 11 th сегмент | |||||||||||||||||
| Vasa дифференциация | 2 пары | 10/11 th to 18 th сегмент | ||||||||||||||||||
| пара простаты | 9025 пара 16/17 -й до 20/21-го сегмента | |||||||||||||||||||
| Дополнительные сальники | 2 пары 902 52 | 17 -й и 19 -й сегмент | ||||||||||||||||||
| Общие простатические протоки | 1 пара | 18 -й сегмент | ||||||||||||||||||
| Генитальная апертура самца | 1 пара | 189 189 |
i.
Тесты:- Семенники — это очень мелкие образования беловатого цвета.
- Есть две пары семенников, они лопастные.
- Каждая пара присутствует в сегменте 10 -й и 11 -й , которые обнаруживаются прикрепленными к задней поверхности межсегментных перегородок 9/10 и 10/11.
- Каждое семенник имеет 4-8 пальцевидных долек, которые содержат округлые клетки в массах, называемых сперматогониями.
- Семенники заключены в семенниковый мешок.
- На молодой стадии дождевого червя семенники полностью развиты, тогда как на взрослой стадии они дегенерируют.
ii.
Яичек:- Мешки яичек — это беловатые, широкие двулопастные мешки, которые покрывают яички.
- Это тонкостенные мешочки, заполненные жидкостью.
- На вентролатеральной стороне брюшного нервного канатика два семенниковых мешка расположены в 10 -м и 11 -м сегментах под желудком.
- В семеннике позади каждого из четырех семенников имеется большая семенная воронка со складчатыми и ресничными краями.
- Мешки семенников соединяются с парой семенных пузырьков.
- Сперматогонии забрасываются в семенники, затем переходят в семенные пузырьки.
iii.
Семенные пузырьки:- Это крупные беловатые сферические образования.
- Они находятся в двух парах, каждая пара расположена в сегменте 11 и 12 сегментов.
- Мешки семенников открываются в семенные пузырьки узким протоком.
- Задние семенные пузырьки крупнее передних.
- Семенные пузырьки 11 -го сегмента находятся заключенными в заднем большом мешочке семенника.
- Семенные пузырьки 12 -го сегмента обнажаются в целомической полости.
- Они расположены вентро-латерально ниже желудка.
- Их также называют перегородочными мешочками, поскольку они развиваются как перегородочные выросты.
- Сперматогонии из семенникового мешка получают семенные пузырьки.
- Семенные пузырьки способствуют питанию сперматозоидов.
- Здесь сперматогонии созревают и образуют сперматозоиды.
iv.
Спермидукальная воронка / семенная воронка:- Они имеют чашеобразную кривизну по конструкции и представлены двумя парами.
- Каждая воронка семявыводящего канала изнутри покрыта ресничками.
- Он расположен под каждым яичком в сегменте 10 -го и 11 -го сегмента .
- Он заключен в тот же мешок яичка.
- Помогает в проведении сперматозоидов.
- После созревания сперматозоиды из семенных пузырьков возвращаются в семенниковый мешок и переходят в дифференцированные сосуды через семявыносящую воронку.
v.
Семявыносящий проток:- Тонкая, длинная, узкая, нитевидная трубчатая структура.
- Сзади каждая семявыносящая воронка ведет к семявыносящему протоку.
- Они находятся в двух парах, и каждая пара расположена по обе стороны от пищеварительного канала.
- Обе пары семявыносящих протоков проходят в непосредственной близости и латеральнее нервного канатика ниже пищеварительного канала на вентральной стенке тела.
- Он простирается от 12 -го до 18 -го сегмента и встречается с протоком предстательной железы в 18 -м сегменте , образуя общий простатический и семенной проток.
- Сперма из семенной воронки собирается семявыносящим протоком и направляется к предстательной железе.
vi.
железы простаты:- Простаты большие, беловатые, плоские твердые, железы неправильной формы.
- Они находятся в паре и расположены по одному с каждой стороны пищеварительного канала в сегментах от 16 -го до 20 -го или 17 -го до 21 st .
- Максимальная часть предстательной железы — это железистая область, а небольшая часть — негландулярная область.
- Толстый изогнутый проток предстательной железы выходит из каждой предстательной железы в сегменте 18 .
- Проток предстательной железы соединяется с двумя семявыносящими протоками своей собственной стороны и образует общий проток простаты и семенного канала.
- На вентральной стороне 18 -го сегмента простатический проток открывается через мужское генитальное отверстие.
- Следовательно, каждое генитальное отверстие имеет три отдельных отверстия, два из которых относятся к семявыносящему каналу и одно — к предстательной железе.
- У дождевых червей секрет простаты полезен для активации сперматозоидов.
- Он также способствует подвижности сперматозоидов.
vii.
Вспомогательные сальники:- Добавочные железы имеют округлую форму и встречаются двумя парами.
- Каждая пара расположена в сегментах 17 -й и 18 -й на брюшной стенке тела на боковых сторонах нервного шнура.
- Эти железы открываются снаружи через набор небольших желез без протоков.
- На двух парах генитальных сосочков, расположенных снаружи по обе стороны от срединно-вентральной линии, эти железы открываются в 17 -м и 19 -м сегментах.
- Считается, что секреция этих желез удерживает двух червей вместе во время процесса совокупления.
- После завершения сперматогенеза образуются хвостатые сперматозоиды.
- Эти сперматозоиды снова попадают в семенники и достигают семявыносящего протока через семенные каналы.
- Затем они выводятся через мужские половые отверстия вместе с секретом предстательной железы.
- Поры половых органов самца расположены попарно и расположены в 18 -м сегменте .
- Женские репродуктивные органы состоят из яичников, яйцевидных трубок, яйцеводов и сперматеек.
- Женский репродуктивный орган включает:
| Структура | Номер | Расположение | |||
| Яичники | 1 пара | thсегмент 1 пара | 13 -я сегмент | ||
| Яйцеводы | 1 пара | 13 -я и 14 -я сегмент | |||
| Генитальная апертура женщины | 9024 th9 один сегмент|||||
| Spermathecae | 4 пары | 6 th , 7 th , 8 th и 9 th сегментов вентролатерально |
i.
Яичники:- Яичники — белые, маленькие, дольчатые структуры, которые встречаются в паре.
- В 13 -м сегменте яичники расположены по одному по обе стороны от брюшного нервного канатика.
- Прилегает к задней части межсегментарной перегородки 12/13 сегментов.
- Каждый яичник состоит из различных пальцевидных выступов, в которых развивающиеся яйцеклетки расположены в ряд, что придает им вид бусинок.
- В каждой доле яичника яйцеклетка проходит несколько стадий своего развития, где зрелые находятся в дистальной части, а незрелые — в проксимальной части.
ii.
Яйцеводы:- Небольшая ресничная воронка яичника с загнутыми краями находится под каждым яичником в 13 -м сегменте .
- Каждая воронка яичника открывается в короткий конический яйцевод.
- Яйцеводы реснитчатые.
- Яйцеводы с обеих сторон соединяются под нервным канатиком.
- Здесь он открывается единственной срединной порой женских половых органов вентрально в сегменте 14 .
- Яйцеклетки после созревания выходят из яичников и попадают в яичниковую воронку.
- Затем он проходит через яйцевод и выводится через поры женских половых органов.
iii.
Сперматека:- Сперматеки — это четыре пары, каждая пара расположена вентро-латерально в 6 -м , 7 -м , 8 -м и 9 -м сегментах.
- Каждая сперматека имеет форму колбы.
- Каждый сперматек состоит из ампулы и шейки (верхняя короткая трубчатая часть).
- основной корпус — ампула.
- На стыке ампулы и шеи обнаружен небольшой мешочек.
- Этот мешок называется дивертикулом.
- Сперматеки также называют семенными вместилищами, поскольку они предназначены для приема сперматозоидов от другого червя во время совокупления и временного хранения сперматозоидов.
- Дивертикул сперматеки Pheretima posthuman хранит сперму, которая попадает сюда после совокупления.
- Но у других видов дождевых червей сперма хранится в ампулах.
- В Pheretima posthuma ampulla способствует питанию сперматозоидов, но у других видов дождевых червей питание обеспечивает дивертикул.
- Семенной проток открывается снаружи небольшими порами, расположенными в бороздках 5/6, 6/7, 7/8 и 8/9 сегментов.
iv.
Пора женских половых органов:- Он однопористый и расположен в сегменте 14 -го .
- Копуляция наблюдалась у нескольких видов дождевых червей.
- Обычно это происходит в сезон дождей с июля по октябрь, в утренние часы перед восходом солнца.
- Во время совокупления два дождевых червя лежат напротив друг друга таким образом, что их вентральные поверхности соприкасаются, а мужские генитальные отверстия одного выходят прямо противоположно семенным отверстиям другого и наоборот.
- Оба дождевых червя получают сперму и секрет простаты во всех своих сперматеках за счет выпуклости, которая возникает из отверстия мужских половых органов.
- Копуляция длится около часа.
- Сперма хранится в сперматеке.
- Оплодотворение происходит только в коконе или яйцеклетке и всегда происходит извне.
- После созревания яичников после совокупления начинается формирование кокона или яйцеклетки.
- В эпидермисе клиторных сегментов присутствуют три разновидности желез: 14 -й , 15 -й и 16 -й сегментов.
- Это слизистые железы, выделяющие слизь для совокупления, железы, выделяющие кокон, производящие стенку кокона, и белковые железы, выделяющие белок, в котором яйца откладываются в коконе.
- В коконе дождевого червя много оплодотворенных яиц.
- Однако только одна из них развивается в эмбрион, тогда как другие оплодотворенные яйца служат питательными / питательными клетками.
- Кокон-секретирующие железы клитора выделяют перепончатый пояс.
- После затвердевания этого пояса происходит отложение альбумина между поясом и стенкой тела.
- Червь начинает отделяться от пояса.
- По мере того, как пояс перемещается по поре женских половых органов, он принимает яйца, а когда он проходит над сперматеками, сперматозоиды выбрасываются в него через поры сперматека.
- Наконец, с переднего конца червя снимается пояс.
- За короткое время эластичность его стенки смыкается с двух концов, образуя кокон или оотеку.
- Несколько коконов образуются после каждого спаривания, поскольку сперматозоиды, присутствующие в сперматеке, не выходят наружу одновременно.
- Коконы имеют овальную структуру, светло-желтого цвета, около 2–2,4 мм в длину и 1,5–2 мм в ширину.
- Оплодотворение происходит внутри кокона, и обычно в коконе находится только один эмбрион.
- Кокон встречается во влажных и прохладных местах, и молодые вылупляются через 2–2,5 месяца.
- Клетки альбумина обеспечивают питание эмбриона.
- Спайность голобластическая, неравномерная.
- Развитие прямое, без личиночной стадии.
- Дождевые черви бывают однодомными, или гермафродитами, или бисексуалами, и протандриями.
- Каждое семенник состоит из 4-8 пальцев, похожих на выступы, и находится внутри семенникового мешка.
- Каждая воронка семенного канала изнутри покрыта ресничками и помогает проводить сперматозоиды.
- Семенные пузырьки у дождевых червей также называют перегородочными мешочками.
- Семенные пузырьки способствуют питанию сперматозоидов.
- У дождевых червей секрет простаты полезен для активации сперматозоидов.
- Каждый яичник беловатого цвета и состоит из яичниковых канальцев.
- Яичники больше семенников.
- Сперматеки имеют форму колбы и находятся по одной паре в каждом из 6 сегментов -го , 7 -го , 8 -го и 9 -го сегментов.
- Каждый сперматек состоит из ампулы и шейки (верхняя короткая трубчатая часть).
- На стыке ампулы и шеи обнаруживается небольшой мешок, называемый дивертикулом.
- Этот дивертикул сперматеек в Pheretima posthuman хранит сперму, которая достигает сюда после совокупления.
- Но у других видов дождевых червей сперма хранится в ампулах.
- В Pheretima posthuma ampulla способствует питанию сперматозоидов, но у других видов дождевых червей питание обеспечивает дивертикул.
- Семенной проток открывается снаружи небольшими порами, расположенными в бороздках 5/6, 6/7, 7/8 и 8/9 сегментов.
- Сперматеки / семеприемники предназначены для приема спермы другого червя во время совокупления и временного хранения сперматозоидов.
- Предполагается, что секреция дополнительных желез помогает удерживать двух дождевых червей вместе во время совокупления.
- Копуляция происходит между двумя дождевыми червями, как правило, ночью в сезон дождей. CJ}?% D = 9vXp # J; lXX0u«x0 Ii6w4 _W [P {) C ۭ ԧ (vAT 뾮 tW8 /
i ~ Jo $ BB 튧 «N; W & v ߖ rpSE
Xwн_ ~ м6 {+ /}] V}> a Om ~ — AW ۯ l Avx h0 @ Ó; 8PG; R0a6
«N-; Ѓagw} / DDDNmTT & DDDDDNy \ gqd إ F» & «] S & cBԧ) [0}? NU0prU * ‘EuD` TI ޛ6 F oS + Z ߯ Պ H # dp 8 DJlY
# iFV, 2pOz & G * фpϒh \ jr> aeƘAυ0 * `RXv`a7A S8Y, pT» tL # pvѰ7.wK_ ߭ a o ߯ d [m)]} К 켿 «N? A9h% ~ ۤ ~ # 9N ~ q +% t |` / 0] aeVu
9G0 Z $ V 鍺 _ (ꎈ> / uKou_ 3, {} zzOzm | 鮻 ޚ _’i}? M *}]} Uj] A դ Uu [_} {Jqe [M582X06VDq @ z5m LPF` ` aa [GppҲa & m / [N (ÍL `_piSlUAxbs !! F8» AXblSL} DdBU «mnt # & k ء i [LS @ ay>»! «» «» «» «+ B» «% Pa
a4 «» «»! «» # ֒ ,, Jt} E «Tv + .̊LeDve1JADEEQ (] = 0PN ۦ 5% TzwVvDApyN
Wo 뮿 P «VuNN8s4ty & 5) TQ́ = r o5 (d _MT` `» H @ 2 f / #: A3 h0Pu $ Q # fu + aCD` «нFq! X; nA @ BӐi
E ذ # `q» V! FhP> Ag? GA.-pd: A | ‘AM n |
7 # s Eѭl
]> ڶ t # [yE ۞ vH6ǃ0H & AFz ~: & m & E {(an [$ tiҤbIe9Al4. $ LBUL6_ O_Z ZI7jm]}
$ o ߫ W_ _ou {dUu_% zz ߺ 0 (x G% | B [gMN) `_.vE 0G «ke ۠ A! O vmG {k} G’j # Dl0A; BwnUNbbq! l0 $ 1P0AA! M1PEcC: bbw? b & i [A7: c
b А «Ямивкж»! ДДДДДДТДД! ah5D ז ZI ׯ KK ը qxz! HȈn (TPcNAr 찥 mGh: gbTT`mvB >; vWauM; _ _ WFn; + K)! a (\ $ f «76 / \ k \ 4» d d1 # A f
A_ \ B $ nAh = 5sGȦJ ٵ «* 21 x` N! WQ` E! ޅ vtXEa
BBC — Земля — Земляные черви занимаются эпическим сексом и вырастают до огромных размеров
На этой неделе земляной червь по имени Дэйв вызвал интерес у британской публики.
Это потому, что Дэйв был измерен в длину 16 дюймов (40 см), что сделало его и ее (мы объясним это позже) самыми длинными дождевыми червями, зарегистрированными в Великобритании.
Но вот фраза, которую вы, возможно, никогда не ожидали прочитать, дождевые черви становятся еще интереснее.
Вот еще пять поразительных фактов.
Дождевые черви вырастают намного больше
Дэйв, червь, возможно, произвел впечатление на Великобританию, включая исследователей, которые измеряли его в Музее естественной истории в Лондоне. Но при 40 сантиметрах он — червяк по сравнению с другими людьми по всему миру. Есть несколько видов, которые могут вырастать более чем на 3 фута (1 м) в длину.
Измерить размер дождевого червя сложно, так как его мягкие тела естественным образом расширяются и сжимаются, а образцы легко непреднамеренно растянуть.
Но самым большим дождевым червем в мире может быть гигантский дождевой червь Гиппсленд в Австралии ( Megascolides australis ), редкий вид, обитающий всего в пяти местах. Согласно Энциклопедии исчезающих видов, особи достоверно достигают 6,6 футов (2 м) в длину при диаметре 4 см. Другой вид, африканский гигантский дождевой червь ( Microchaetus rappi ), также, как сообщается, достигает длины 6.6 футов (2 м) или более.
Гигантские дождевые черви создают загадочные холмы
Дождевые черви известны своей подвижностью почвы. Но в Южной Америке один вид был признан создателем красивой и загадочной серии курганов.
Суралес — это ландшафт зеленых курганов и глубоких ям, организованных в замысловатые и правильные узоры. Они простираются на многие квадратные километры, но эти ошеломляющие насыпи были построены скромными дождевыми червями, которые вырастали более чем на 3 фута (1 м) в длину.
Даже то, как дождевые черви строят холмы, немного странно. На протяжении десятилетий черви Andiorrhinus придерживались жестких пищевых привычек. Они едят в затопленных ямах и всегда какают в одном и том же месте.
Со временем их какашки накапливаются в большие насыпи до 16 футов (5 м) в поперечнике и 7 футов (2 м) в высоту. Это открытие было опубликовано учеными только в мае 2016 года.
У земляных червей эпический секс
Что касается сексуальной выносливости, у дождевых червей другие виды, включая людей, легко побеждают.Обычный дождевой червь ( Lumbricus terrestris ) совокупляется более трех часов за раз.
Исследование 1997 года показало, что дождевые черви ухаживают друг за другом, посещая норы друг друга, иногда до 17 раз.
Если это не проверяет их силу духа и запасы энергии в достаточной степени, каждое фактическое совокупление длится 69-200 минут. Ученые знают это, потому что они засняли действие дождевых червей.
Воспитание затруднено
Дождевые черви — гермафродиты, у каждой особи есть как мужские, так и женские половые органы.Когда они спариваются, они соревнуются за оплодотворение друг друга спермой и оплодотворение яйцеклеток другого. Это означает, что дождевой червь Дейв мог быть и матерью, и отцом для разных потомков, передавая каждому половину своих генов.
Но осложнения продолжаются.
В 2013 году ученые впервые провели тесты на отцовство дождевых червей. Они обнаружили, что у дождевых червей часто бывает несколько партнеров, и первый и третий дождевые черви, вступившие в половую связь с партнером, с наибольшей вероятностью дадут потомство.Те, кто идут вторыми, теряют, возможно, потому, что их сперматозоиды должны стоять в очереди, чтобы достичь резервуара для хранения, который был заполнен первым партнером, и смывается спермой, введенной третьим.
Более того, в то время как у некоторых дождевых червей бывают продолжительные приступы секса, другие виды, похоже, полностью покончили с сексом.
Гениталии некоторых тайваньских горных дождевых червей ( Amynthas catenus ) дегенерировали. Вместо этого они размножаются через непорочное зачатие или «партеногенез».Это означает, что они передают 100% своих генов потомству, производя клоны самих себя. Это делает их отцом и матерью каждого детеныша червяка.
Подобно коровам, дождевые черви образуют стада
И они принимают групповые решения. Согласно исследованию, опубликованному в 2010 году, дождевые черви используют прикосновения, чтобы общаться и влиять на поведение друг друга.
Таким образом черви коллективно решают двигаться в одном направлении в составе одного стада. Поразительное поведение, обнаруженное у дождевого червя Eisenia fetida , является первым случаем, когда любой кольчатый червь может образовывать активные стада.
Открытие предполагает, что дождевые черви — социальные животные, и их групповое поведение похоже на стадо или роение других видов.
Итак, в следующий раз, когда вы возьмете червя и бросите его в компостную кучу, запомните это. Вы можете оторвать его от друзей.
Следуйте за Мэттом Уокером в Twitter: @byMJWalker.
Присоединяйтесь к более чем шести миллионам поклонников BBC Earth, поставив нам лайк на Facebook или подписавшись на нас в Twitter и Instagram .
Если вам понравился этот рассказ, подпишитесь на еженедельную рассылку новостей bbc.com под названием «Если вы прочитаете только 6 статей на этой неделе». Тщательно подобранная подборка историй из BBC Future, Earth, Culture, Capital, Travel и Autos, которые доставляются вам на почту каждую пятницу.
Земляных червей — Друзья на заднем дворе
Как выглядят дождевые черви?
В Австралии насчитывается более 1000 видов местных червей и около 80 интродуцированных видов, которые также являются полезными.Не все они маленькие и коричневые — некоторые аборигенные австралийские черви огромны. «Гиппслендский гигант» занесен в Книгу рекордов Гиннеса по росту до 3 метров в длину. Вид, обитающий на северо-востоке Нового Южного Уэльса, часто вырастает более 150 см и толщиной с садовый шланг. Но вряд ли вы встретите одного из этих монстров у себя на заднем дворе.
Где водятся дождевые черви?
Дождевые черви, обитающие по всей Австралии, — отличные друзья в вашем саду.Они возвращают в почву питательные вещества из органических веществ, таких как опавшие листья, кожура овощей, фруктовые обрезки, обрезки волос и даже старая бумага. Эти питательные вещества важны для растений и значительно обогатят почву на вашем заднем дворе.
Быстрые факты:
- У червей есть мужские и женские органы, но для размножения им нужен еще один червь. Они откладывают яйца, которые вылупляются примерно через три недели.
- У дождевых червей нет легких, они дышат через кожу!
Земляные черви
В Австралии насчитывается более 1000 видов местных червей и около 80 интродуцированных видов, которые также являются полезными.
Дождевые черви — отличные друзья в вашем саду. Они возвращают в почву питательные вещества из органических веществ, таких как опавшие листья, кожура овощей, фруктовые обрезки, обрезки волос и даже старая бумага. Эти питательные вещества важны для растений и значительно обогатят почву на вашем заднем дворе.
Не все они маленькие и коричневые — некоторые аборигенные австралийские черви огромны. В Книге рекордов Гиннеса зафиксирован рост «гиганта Гиппсленда» до 3 метров. Вид, обитающий на северо-востоке Нового Южного Уэльса, часто вырастает более 150 см и толщиной с садовый шланг.Но вряд ли вы встретите одного из этих монстров у себя на заднем дворе.
Дождевые черви могут ежедневно съедать до половины своего веса органическими веществами. Проходя туннели и роясь под землей, дождевые черви аэрируют вашу почву, делая ее менее компактной и облегчая проникновение воды к корням растений.
У червей есть веская причина оставаться под землей — они не только должны оставаться влажными, но и являются любимой пищей многих птиц.
Избегайте использования химикатов или пестицидов в вашем саду, так как они могут попасть в почву и вызвать заболевание ваших червей.Если вы недавно гнали своего питомца, соберите весь помет в саду и выбросьте его в мусорное ведро, так как эти химические вещества могут убить дождевых червей.
Помещая органические отходы, такие как кожура фруктов и овощей, обрезки и даже измельченная бумага, на компостную или червячную ферму, вы сокращаете количество мусора, отправляемого на свалку.
Будьте приятелем на заднем дворе
Любовь дождевых червей:
- Мертвые органические вещества, такие как опавшие листья, кожура овощей и фруктовые обрезки, которые они собирают под землей и едят.
- Мульча и почвопокровное растение, которые сохраняют почву прохладной и влажной.
- Ночное время, когда они выходят из почвы, чтобы схватить листья и объедки, вытащить под землю и поесть. Днем они остаются под землей, чтобы не высыхать и избегать хищников.
- Туннелирование для передвижения под землей. Это также аэрирует вашу почву.
А им не нравится:
- Сильные ароматы, такие как цитрусовые, ананас, перец чили, лук, чеснок или лук-шалот.
- Мясо, курица или рыба, как дождевые черви, относятся к вегетарианцам.
- Химические вещества, масло или пестициды, вызывающие заболевание.
Будьте другом земляным червям
Попробуйте:
- Сделайте компостную кучу в своем саду.
- Прикрывайте червей, которых вы обнаружите, копая в саду. Они не любят, когда их разоблачают.
- Мульчируйте грядки, вырастите почвопокровное растение или позвольте опаду из листьев стать вашей мульчей. Это обеспечит червям достаточное количество пищи и сохранит почву влажной.
- Соберите все овощные кожуры, фруктовые обрезки, старую измельченную бумагу и положите их в сад, чтобы червяки поели.
- Создайте червячную ферму и приготовьте сок червя (жидкое удобрение), который отлично подходит для ваших растений. В хороших условиях черви размножаются каждые 7-10 дней, для вылупления яиц требуется около 21 дня, а через 2-3 месяца новые черви готовы к размножению. Население будет удваиваться каждые 2-3 месяца и будет съедать все ваши отходы и органические материалы.
Избегать:
- Использование химикатов, пестицидов или инсектицидов в вашем саду.
- Перелив вашего сада, поскольку черви любят влажную, но не очень влажную почву.
- Помещение мяса, молочных продуктов, сладких продуктов, острых овощей или цитрусовых в огород, поскольку черви не любят их есть и могут привлечь других насекомых и крыс.
Добавить комментарий
Комментарий добавить легко